獣脚類を中心とした恐竜イラストサイト
肉食の系譜




アンズー

Copyright Lamanna et al.
アンズーは、白亜紀後期マーストリヒト期(ヘル・クリーク層)に米国ノースダコタおよびサウスダコタ州に生息した大型のオヴィラプトロサウルス類で、2014年に記載された。保存の良い3体分の部分骨格が発見されており、全部合わせると体のほとんどの骨格がそろっている。これまで断片的だった北アメリカのオヴィラプトロサウルス類(カエナグナトゥス科)の形態や系統関係などを解明するのに役立つと期待されている。
大部分のオヴィラプトロサウルス類はアジア、特にモンゴルと中国で発見されている。白亜紀後期のアジア産オヴィラプトロサウルス類のほとんどは、単系のオヴィラプトル科Oviraptoridaeをなす。一方、北アメリカからもオヴィラプトロサウルス類の化石が長く知られているが、残念なことに北アメリカ産のオヴィラプトロサウルス類の標本は非常に不完全なため、その解剖学的形態や分類、系統関係の詳細は明らかでなかった。多くの研究者は、白亜紀後期の北アメリカ産オヴィラプトロサウルス類は、すべて単系のカエナグナトゥス科に属すると考えているが、最近のいくつかの研究ではカエナグナトゥス科は多系群であると考えられている。さらに、多くの研究者がこれらの恐竜の生態・習性について推測しているが、それぞれがまちまちな結論を出している。
系統解析の結果、オヴィラプトロサウルス類の中のいくつかのグループ(カウディプテリクス科 Caudipterygidae、 カエナグナトゥス上科 Caenagnathoidea、 オヴィラプトル科 Oviraptoridae、カエナグナトゥス科 Caenagnathidae)が単系であることが支持された。アンズーは派生的なカエナグナトゥス科のメンバーで、カエナグナトゥスと姉妹群と考えられた。また、白亜紀前期のミクロヴェナトルとギガントラプトルは基盤的なカエナグナトゥス類と位置づけられた。
アンズーの発見によって初めて、カエナグナトゥス科のオヴィラプトロサウルス類のほとんど完全な骨格形態が明らかになってきた。アンズーは推定全長3.5 m、腰の高さ1.5 m、体重200-300 kg もあり、ギガントラプトルに次いで最も大型のオヴィラプトロサウルス類の一つである。カエナグナタシア(推定体重5 kg)のような非常に小型の種類もいることを考え合わせると、カエナグナトゥス科は獣脚類の中でも、体のサイズが非常に広範囲にわたるグループということになる。これを裏付けるためにはカエナグナトゥス類の成長や個々の標本の成長段階の研究が必要になってくるという。
アンズーの頭骨は丈が高く幅が狭く、ヒクイドリのような高いトサカがある。意外なことに、頬骨はオヴィラプトロサウルス類以外の獣脚類のものと似ている。上下の顎には歯がなく、左右が癒合した歯骨の咬合面には特徴的な稜と溝の列が並んでいる。首は長く12個の頸椎からなり、後方にいくにつれて幅が広くなっている。尾椎は含気性が発達していて、後端には特殊化した椎骨が並び、全体として尾端骨状の構造をなしている。胸骨板はオヴィラプトル科のものとよく似ている。手は比較的大きく、末節骨の基部には他の多くのオヴィラプトロサウルス類よりも顕著な唇状の部分lipがある。手の第2指と第3指はアンズーでは不完全であるが、他の手がよく保存されたカエナグナトゥス類(キロステノテス、エルミサウルス)と同様におそらく長かったと思われる。後肢の骨は細長く、脛骨が大腿骨よりもかなり長い。足もアンズーでは不完全であるが、見つかっている部分から他のカエナグナトゥス類と同様に足の指が長かったと考えられる。
顎に歯がなく角質で覆われたくちばしをもつオヴィラプトロサウルス類の食性については、さまざまな推測がなされてきた。最初の仮説はオヴィラプトルが卵泥棒というもので、その後オヴィラプトル類が自分自身の卵を守っていることがわかり、これ自体は否定された。しかしオヴィラプトル類が他の脊椎動物の卵を食べた可能性は否定されていない。Currie et al.は、オヴィラプトル類の上顎骨の口蓋部分にある歯のような突起を、卵を食べるヘビが卵殻を割るための椎骨の突起に例えて、オヴィラプトロサウルス類は卵や小型の脊椎動物を食べただろうと論じている。
Barsboldはオヴィラプトル類の頑丈な顎は貝を割るために用いられたと提唱したが、この説は解剖学的、生物力学的、古生態学的根拠から強く否定されている。Cracraft と Funston and Currieは、カエナグナトゥス類の下顎と、植物食の単弓類ディキノドン類の下顎の類似性に注目した。カエナグナトゥス類とディキノドン類の顎関節の構造は、下顎を前後に大きく動かせたことを示しており、それにより植物を上下のくちばしのある顎で刈り取るのに役立ったかもしれない。とくにFunston and Currieは、カンパニア期のカエナグナトゥス類の下顎とディキノドン類の下顎を詳細に比較し、ディキノドン類で植物食への適応と考えられている5つの特徴のうち4つまでを、カエナグナトゥス類はみたすとしている。面白いことに、アンズーとギガントラプトルではさらに5番目の特徴までみたしている。それは歯骨の側面にあるフランジである。この構造は、ディキノドン類では前方にある内転筋の付着部であると考えられており、この2種のカエナグナトゥス類でも同様に機能したかもしれない。
オヴィラプトロサウルス類が植物食だったという最も有力な証拠は、歯のある原始的な種類でのみ知られている。カウディプテリクスのいくつかの標本には、胃石が保存されていた。またインキシヴォサウルスの前上顎骨の歯は齧歯類の切歯に似ている。さらに、最近記載された基盤的なオヴィラプトロサウルス類、ニンギュアンサウルスの体腔内には多数の卵形の構造物が保存されており、種子かもしれないと考えられている。おそらく基盤的なオヴィラプトロサウルス類は完全にあるいは主に植物食であったが、進化した種類では角質のくちばしが発達して、小動物や卵など広範囲の食物を食べるようになったのではないか、としている。
カエナグナトゥス類の習性については、四肢の骨格の特徴からもいろいろと考察されている。Currie and Russellは、比較的長い後肢と指の長い広がった足先から、カエナグナトゥス類は渉禽類のように浅瀬に入り、水中の無脊椎動物を捕食したのではないかと考えた。彼らはキロステノテスの細長い第3指は、川底や木の割れ目から小動物をかき出すのに役立っただろう、ともいっている。
カエナグナトゥス類の化石が発見される地層の堆積環境が、生息地を推定する手がかりになるかもしれない。カエナグナトゥス類とオヴィラプトル類では、古生態学的に明らかな違いがあることが、長い間注目されてきた。オヴィラプトル類の化石のほとんどは、乾燥あるいは半乾燥気候の環境とされる地層から発見されているのに対し、多くのカエナグナトゥス類の化石はより湿潤な状態で堆積したと考えられる河川堆積物から見つかっている。よってカエナグナトゥス類は、近縁のオヴィラプトル類よりも水の豊富な、湿潤な環境に適応していたと考えられている。
アンズーのより完全な2つの標本は、川岸の堆積物と考えられるシルト状泥岩から見つかっている。1つの断片的な標本は河床の堆積物から見つかっている。ヘル・クリーク層の別のカエナグナトゥス類の部分骨格は、やはり有機物の豊富な泥岩から発見されている。これは死体が流された可能性もあるものの、真の古生態を反映しているかもしれない。つまりヘル・クリーク層のような古環境では、カエナグナトゥス類は氾濫原のような湿地を好んだ可能性がある。
結局、アンズーや近縁のカエナグナトゥス類は、当時の北アメリカの沿岸部の平野で湿潤な環境を好み、植物や小動物、卵など何でも食べる雑食動物だったのではないか、としている。
参考文献
Lamanna MC, Sues H-D, Schachner ER, Lyson TR (2014) A New Large-Bodied Oviraptorosaurian Theropod Dinosaur from the Latest Cretaceous of Western North America. PLoS ONE 9(3): e92022. doi:10.1371/journal.pone.0092022

コメント ( 1 ) | Trackback ( 0 )
アリオラムス・アルタイ2014


大きい画像
アリオラムス特集の記念イラスト。また何か面白いことを見つけたら書きます。
コメント ( 1 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(10)一応完
Brusatte et al. (2012) は、最後に「新しいボディプランと生態学的習性」という項目で考察しているが、あまり目新しいことは言っていない。
アリオラムスの特徴的に長い頭骨は、タルボサウルスやティラノサウルスのような同時代の他のティラノサウルス類とは異なる生活様式をとっていたことを示唆する。アリオラムスの特徴としては長い吻のほか、全般にきゃしゃな頭骨の骨、互いに入り組んだinterlocking縫合線がないこと、比較的薄い歯、眼窩の上にがっしりした涙骨/後眼窩骨の「ひさし」lacrimal-postorbital barがないこと、顎の筋肉の付着面が比較的小さいこと、体格が小さいこと、頭骨および胴体の骨で含気性が非常に発達していることなどがある。これらの特徴のうちいくつかは、このホロタイプが亜成体であることに関係していると考えられ、どれとどれが成体になっても維持される形質かは予測が難しいという。
しかし、これまで見つかっているアリオラムスの標本では、大型ティラノサウルス類の成体に特徴的な「噛み潰す」‘‘puncture-pull’’捕食様式はとれなかったことは明らかである。他の大型ティラノサウルス類では、強力な筋肉、太い歯、頭骨の入り組んだ縫合線、圧力を分散させる眼窩の「ひさし」などにより、骨を砕くような強い力で咬むことが可能になっている。少なくとも幼体から亜成体のアリオラムスでは、これとは別の捕食様式をとり、つまりより小型の獲物を狙い、咬む力よりもスピードを重視したのだろうとしている。これはおそらくタルボサウルスやティラノサウルスの幼体にもあてはまることだろう。完全な成体のアリオラムスが「噛み潰す」捕食様式をとれたのかどうかは、成体の化石が発見されて精密な生体力学的解析がなされるのを待たなければならない。
アリオラムス・アルタイのホロタイプと同じ地域からタルボサウルスの化石も見つかっているので、両者は共存していたはずである。このように2種類の大型捕食者が生息していた状況は、ダスプレトサウルスとゴルゴサウルスが共存していたカンパニア期の北アメリカと似ている。これらはニッチ分割niche partitioningによって共存できたと考えられており、タルボサウルスとアリオラムスも異なる獲物を捕食することですみわけていたのだろう、という。(このあたりはもう少し具体的な考察を期待したのであるが、無難なことしか書いていない。)ジュラ紀後期の北アメリカの大型獣脚類相(アロサウルス、ケラトサウルス、トルボサウルス)などと比べて、白亜紀後期では大型捕食者がいずれもティラノサウルス類で構成され、他のグループの獣脚類がいないのが特徴的であるという。
大型ハドロサウルス類やよろい竜はタルボサウルスに任せて、アリオラムスは中型から小型の動物を狙ったのだろう。大型の恐竜がほとんど生息せず、オヴィラプトル類やトロオドン類や哺乳類ばかりが多い地域があれば、アリオラムスにとっては都合がよかったのかもしれない。いずれにしてもタルボサウルスの幼体とは競合すると思われる。
コメント ( 0 ) | Trackback ( 0 )
たんば恐竜塾


平山先生と徳川さんを講師に迎えた「たんば恐竜塾」に参加するため、久しぶりにちーたんの館へ行ってきました。ここを訪れるのは2回目ですが、最初は開館まもなくだったのでガストニアは初めてです。
まず平山先生の解説つきで丹波竜についての見学。その後発掘現場での石割りのほか、エアーチゼルを使ったクリーニング体験など貴重な経験ができました。平山先生の講演は、(最近出版された著作にもある)竜脚類の首とステゴサウルスの食性のお話でしたが、夜の部がハードで。。。徳川さんのドイツ紀行を含め、単弓類から獣脚類から、オドントケリスまで。中には知りたくなかったような「不都合な真実」まで、裏話が満載でした。
2日目は骨格復元ワークショップで、午後は篠山市民センターに移動して、ササヤマミロスやトカゲ化石などについての公開講座と、大変充実した週末になりました。企画・運営に携わった丹波市恐竜課の皆さん、ありがとうございました。(平山先生はご飯が食べたかったようですよ。)
忘れてはいけないのが、最近発売された丹波竜復元模型です。竜脚類展もあるようなので行かれる方はどうぞ。関西の方は福井のシンポジウムなどにも行きやすくていいですね。
コメント ( 0 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(9)
Brusatte et al. (2012) は、アリオラムスの長い頭骨がどのように生じたかについてDiscussionで考察している。アリオラムスの特徴的な長い頭骨は、個体発生の過程で起きた2つの変化によって生じただろうとしている。
まず一つは、個体発生の過程で頭骨の丈が高くなるような成長が抑制され、成長しても幼体のような丈の低い頭骨と下顎を保つようになった。これはプロケラトサウルス、グァンロン、ディロングのような基盤的ティラノサウロイドにみられる原始状態への逆戻りかもしれない。派生的なティラノサウルス類では幼体に比べてはるかに頭骨の丈が高くなる。このことは上顎骨の本体の丈や眼窩の形など多くの形質に表れる。
次にもう一つ別の要因によって、眼窩から側頭部の部分orbitotemporal regionはそのままで、吻の部分だけが長くなるような変化が起こった。アリオラムスの頭骨では、涙骨の前方突起や鼻骨の前頭骨突起が他のティラノサウルス類に比べて長くなっている。つまり、前眼窩窓の周辺の骨が特に長くなっているのである。この部分は、頭骨のより前方部分(吻の前端)や後方部分(眼窩から側頭部)に比べて、骨も少なく複雑な軟組織構造(脳、感覚器、気嚢)もあまりない。よって機能をそこなうことなく、変化が生じることができた。基盤的なティラノサウロイドであるシオングァンロンも、同様にして長い頭骨を発達させたのかもしれない。
アリオラムスでは前眼窩窓の領域が単純に引き延ばされるようにして吻が延長しているので、主要な軟組織構造としては気道に影響するくらいであったと思われる。この変化は口蓋骨にも及んでおり、口蓋骨の鋤骨翼状骨突起vomeropterygoid process は涙骨から離れて前方に位置している。より吻が短いゴルゴサウルスやティラノサウルスでは、鋤骨翼状骨突起はもっと涙骨と近い位置にある。またアリオラムスでは前眼窩窓の前縁は口蓋骨よりもかなり前方にある。一方、吻が短いティラノサウルス類では、口蓋骨の鋤骨翼状骨突起の先端が前眼窩窓の前縁に近づいている。
アリオラムスでは上顎骨と歯骨の歯列の最後の歯が、前眼窩窓の前半部の下にあり、頬骨のずっと前方にある。このことも前眼窩窓の部分が長くなっているという考えと一致する。それに対して吻が短いティラノサウルス類では、最後の歯は前眼窩窓の中央の下にあり、頬骨と非常に近いという。
派生的なティラノサウルス類の形態は比較的保守的であるが、変わらないわけではない。アリオラムスとシオングァンロンの頭骨は、機能に悪影響を与えることなく、大きな形態変化を達成することができることを示している。同様に白亜紀後期のテラトフォネウスは、派生的なティラノサウルス類が吻を短くすることもできることを示した。ティラノサウルス類の頭骨の形態は、従来考えられていたよりも多様であることがわかってきた、としている。
Brusatte et al. (2012) はアリオラムスをティラノサウルス科の中と考えているので(系統解析は2010の総説)、このような考察になる。リスロナクスの論文のようにもう少し基盤的でティラノサウルス科の外と考えると、多少違う論じ方になるだろう。当然ながらこのモノグラフを読んでいると、やはりティラノサウルス科の中のような気がしてくる。もう少し原始的な種類がどういう形態かがわかってくると、真実が解明されてくるだろう。
ホッキョクグマは分子系統上はヒグマと非常に近縁だが、短期間に大きく形態が変化したという話があった(現在どうなっているか知らないが)。派生的なティラノサウルス類が幼形進化などで大きく形態変化した場合に、形態情報のみの解析でうまく扱えるのだろうか。かなり進化した種類で吻が伸びた場合と、やや原始的な種類で吻が長かった場合とで区別できるのだろうか。その場合、吻の長さ以外の形質が重要なのだろう。
コメント ( 4 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(8)


それでは、Gaston社の「復元」頭骨レプリカ、つまり全身骨格の頭はどうなのだろうか。「産状」と「復元」ではなくて、「復元」ver.1とver.2ならば、実はver.2の方が正確だったりしないのだろうか。
そこで2012年の写真を見ると‥‥orzである。少なくともmaxillary fenestraと後眼窩骨については、やはり残念なことになっている。maxillary fenestraは前眼窩窩の前縁に近づけられている。後眼窩骨の角状突起は「産状」ほど盛られてはいないかもしれないが、眼窩下突起は明らかに作られている。
こんなことで私のアリオラムス愛は揺るがないが、惜しいことである。世の中には知らない方が幸せなこともある例かもしれない。それにしても、すばらしい頭骨ですね。
コメント ( 3 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(7)


口蓋骨は、左右ともほとんど完全に保存されている。口蓋骨は四放射状で、4つの突起からなっている。上顎骨突起maxillary process、頬骨突起jugal process、鋤骨翼状骨突起vomeropterygoid process、内側突起medial processである。口蓋骨は頭骨内部の骨であるが、一部は前眼窩窓を通して見ることができる。
鋤骨翼状骨突起vomeropterygoid processは背方にのびている突起で、上方突起ascending process、背内側突起dorsomedial processなど他の名前でもよばれる。アリオラムス・アルタイでは、鋤骨翼状骨突起が前後に拡大しており、前方突起anterior projection と後方突起posterior projectionがある。後方突起は短いが、前方突起は非常に長く伸びて、先細りの三角形をしている。このような形態は他のティラノサウルス類と比べて特徴的である。
タルボサウルスとティラノサウルスでは鋤骨翼状骨突起の上端は拡大しておらず、長方形をしている(前方突起と後方突起はあるが、短い)。アルバートサウルスとダスプレトサウルスでは上端が拡大しているが、尖った先端に向かって先細りになってはいない。アリオラムス・アルタイの鋤骨翼状骨突起の形は、アパラチオサウルスと似ている。
頬骨突起の基部付近に、大きく深い含気孔である口蓋骨窩palatine recessがある。アリオラムス・アルタイでは、口蓋骨窩が鋤骨翼状骨突起の後端よりもずっと後方までのびている。このことはBrusatte et al. (2009) ではアリオラムス・アルタイの固有形質とされたが、ダスプレトサウルスの少なくとも一つの標本にはみられる。しかしそれ以外のティラノサウルス類にはみられない。他のティラノサウルス類では、口蓋骨窩は鋤骨翼状骨突起の後端よりも前方にあるか、ちょうど後端の位置にある。

ティラノサウルス(スタン)の右の口蓋骨。長方形というのは四角いテーブル状ということかと思われる。左の口蓋骨では鋤骨翼状骨突起が欠けているらしい。
コメント ( 0 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(6)
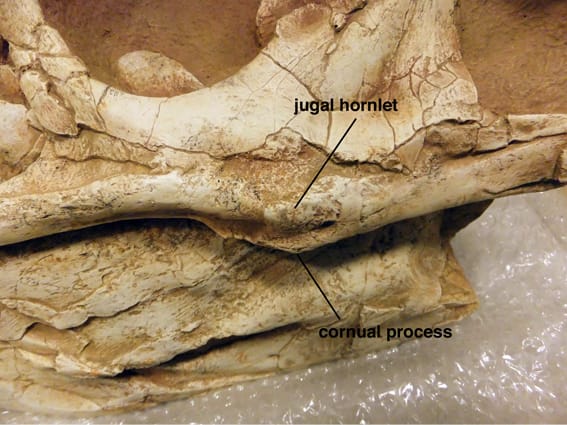
頬骨は左側しか見つかっていないが、左の頬骨はほとんど完全に保存されている。頬骨には、上顎骨および涙骨と結合する前方突起anterior ramus、後眼窩骨と結合する背方突起dorsal ramus、方形頬骨と結合する後方突起posterior ramusがある。
頬骨の外側面は全般に平滑であるが、前方突起から眼窩の下にかけて粗い部分がある。アリオラムス・アルタイの頬骨で最も顕著な特徴は、眼窩の後縁の真下で側方に突出した骨の突起で、これを頬骨小角jugal hornlet という。頬骨小角jugal hornletは、他のどのティラノサウルス類にもみられないもので、アリオラムス・アルタイの固有の特徴である。ただしアリオラムス・レモトゥスについてははっきりしない。Kurzanov (1976)はこの部分に「角質で覆われていた可能性のある、結節のある表面」という記述を残している。
ティラノサウルス類全般には、眼窩の下方にこれとは別の構造がある。頬骨の腹側縁にある突起で、(頬骨の)角状突起cornual process (of jugal) という。角状突起はアリオラムス・アルタイにもあり、はっきりしている。これはビスタヒエヴェルソル、アルバートサウルス、ゴルゴサウルスなどと似ている。この突起はティラノサウルスの成体でははっきりしない。一方、ダスプレトサウルス成体では最も顕著であるという。

頬骨は、他のティラノサウルス類と同様に高度に含気性である。前方突起の外側面で、前眼窩窩の後腹側部分にあたるところに、大きな含気窩pneumatic recessがある。アリオラムス・アルタイでは、2つの穴がつながったような、ピーナッツ形をしている。つまり含気窩の前方ポケット(app, anterior pocket of pneumatic recess)と後方ポケット(ppp, posterior pocket of pneumatic recess)という2つの穴からなっている。他のティラノサウルス類では、多くの場合、後方ポケットしかない。前方ポケットが存在する場合でも、それは小さく後方ポケットとつながっていない。アリオラムス・アルタイの後方ポケットの形は、ゴルゴサウルスのものとよく似ており、涙形teardrop-shapedで45度傾いている。一方、ダスプレトサウルス、タルボサウルス、ティラノサウルスの含気窩は一般に小さく、水平または垂直であるという。またタルボサウルスの含気窩は、より円形に近いという。

タルボサウルスでは含気窩が1個で円形に近い
コメント ( 1 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(5)


大きい画像
今回、最もがっかりしたのは後眼窩骨である。まず図をご覧下さい。
後眼窩骨はT字形で、前方突起、後方突起、腹方突起からなる。前方突起の腹側縁、つまり眼窩の後背方部分の上にあたるところに、最も顕著な粗面がある。アリオラムス・アルタイでは、ここにごつごつした稜があり、細かい溝と結節からなっている。この稜は、ティラノサウルス科にみられる角状突起cornual process (of postorbital) と相同なものである。
後眼窩骨の角状突起は、ディロング、グァンロン、シオングァンロン、ラプトレックスのような基盤的なティラノサウロイドにはない。一方、ティラノサウルス科とビスタヒエヴェルソルでは、眼窩の後背方部の上にごつごつした隆起がある。多くの種類ではアリオラムスよりも大きく膨らんでいて、角といわれるような突出した構造に発達している。これはアルバートサウルス、ビスタヒエヴェルソル、ゴルゴサウルスにみられ、タルボサウルスとティラノサウルスで特によく発達している。最も発達した「角」はダスプレトサウルス成体にみられるという。
アリオラムス・アルタイの角状突起は低い稜状で、ダスプレトサウルスの幼体と最もよく似ている。Kurzanov (1976) の図と記述によるとアリオラムス・レモトゥスも同様で、はっきりと膨らんではいないという。
Gaston社の「産状」頭骨では、あれ?確かにこの位置に角状突起があるのはいいが、膨らみすぎである。こんなに取っ手状に張り出してはいないはずである。このレプリカでは角状突起の後方にはっきりと崖ができており、ホロタイプの写真とは違いすぎる。「眉メイク」にもほどがある。
アリオラムス・アルタイのホロタイプの腹方突起には、眼窩の中に突出した眼窩下突起suborbital processはない。これはゴルゴサウルス、ダスプレトサウルス、ティラノサウルスの幼体と同様である。一方、多くのティラノサウルス科には眼窩下突起がみられる。Brochu (2003) は、眼窩下突起はタルボサウルスとティラノサウルスにみられ、ダスプレトサウルスでは萌芽的状態としている。またアルバートサウルスとゴルゴサウルスには眼窩下突起はないとしているが、Currie (2003) によると大型の個体にはある。眼窩下突起の大きさや形は成長段階により変化するもので、ティラノサウルス亜科でもアルバートサウルス亜科でも幼体にはない。
Gaston社の「産状」頭骨では、あれ?なんであるの?しかも形がどうも不自然である。腹方突起の先端が二股に分かれていて、そこに妙な楕円形の部分がある。これが本来の腹方突起の先端であるようにみえる。つまり、無理矢理、眼窩下突起を作ったようにみえる。眉メイクだけではなく、「つけまつげ」もつけられている。
要するにこの「産状」頭骨レプリカは、ティラノ・タルボ風にメイクされている。(激怒)この方がなんとなく見栄えがすると思ったのだろうか。

タルボサウルス(亜成体か若い成体と思われる)では角状突起が発達し、眼窩下突起もある。
コメント ( 2 ) | Trackback ( 0 )
アリオラムスの頭骨を観察しよう(4)


大きい画像
涙骨は左右とも見つかっているが、左の涙骨がより完全に保存されている。涙骨は、主に前方突起anterior ramusと腹方突起ventral ramusからなる。アリオラムス・アルタイでは前方突起と腹方突起が鋭角をなし、涙骨は数字の7の形をしている。これはダスプレトサウルス、タルボサウルス、ティラノサウルスの成体と同様である。一方、アルバートサウルス、アパラチオサウルス、ビスタヒエヴェルソル、ゴルゴサウルス、ラプトレックス、およびダスプレトサウルス、タルボサウルス、ティラノサウルスの幼体では、前方突起と腹方突起が直角をなし、逆L字形に近い。
アリオラムス・アルタイでは前方突起が特に長い。前方突起/腹方突起の比率はアリオラムス・アルタイが1.40、ティラノサウルスが0.90である。
前方突起も腹方突起も、外側板lateral laminaと内側板medial laminaという2枚の板が重なった形をしている。前方突起の外側板と内側板の間には、いくつかの含気孔がある。アリオラムス・アルタイのホロタイプでは、アルバートサウルスなどの幼体と同様に、外側板はあまり発達していない。
腹方突起の外側板の表面は粗面になっている。外側板の前縁の方が後縁よりも盛り上がっていて、その間に眼窩窩orbital fossaという凹みがある。また腹方突起を横切るように前後に走る、数本の弧状の線条arcuate striationがある。
涙骨で最も顕著な隆起が、前方突起と腹方突起が交わる部分の背側にある角状突起である。アリオラムス・アルタイの角状突起は、はっきりした円錐形に近い突起で、その根元には一連のいぼ状突起ovoid excresences (骨乳頭bone papillae)があって境界をなしている。
(Gaston社のレプリカの涙骨は、大体正確にできているが、精度の問題はある。線条arcuate striationはホロタイプではもっと多く腹側にもあるのだが、レプリカでは写真の2、3本しかはっきりとは見えない。角状突起の境界にあるといういぼ状突起は、心眼でしか見えない。しかしまあ頭骨全体のレプリカだから、このくらいは精度的に仕方ないかもしれない。ただ前方突起の外側板は、ホロタイプに比べて垂れ下がりすぎている。ここは「成体化」されている。)
Brusatte et al. (2012) は、角状突起について1ページ以上費やして詳述している。ティラノサウルス類の角状突起は、その形、大きさ、位置などが種類によっても成長過程によっても変化する。
アリオラムス・アルタイの角状突起は、涙骨の背側縁の残りの部分からはっきりと突出した構造であるといっている。涙骨の背側縁のうち、いぼ状突起の境界で囲まれた内側だけが角状突起という意味だろう。成体のアルバートサウルス、ビスタヒエヴェルソル、ゴルゴサウルス、ダスプレトサウルス、テラトフォネウスはいずれも顕著な角状突起をもつが、これらはより大きく、前後に広く、涙骨の背側縁になめらかにつながっているという。しかし、アルバートサウルスをはじめダスプレトサウルス、タルボサウルス、ティラノサウルスなどの幼体には共通して、小さくはっきりした角錐状の角があるとも述べているので、このあたりは種の特徴と成長段階の特徴(亜成体であるがゆえの)がうまく区別されていないように思える。
それでも著者らは、円錐形の角状突起が涙骨の背側縁から急に立ち上がった形をしていることをアリオラムス・アルタイの固有の形質と考えている。一方、ゴルゴサウルスなどの亜成体では、角状突起の前方は急に突出しているが、後方は涙骨の背側縁になめらかにつながっているという。要するに特徴は角状突起の後方のカーブということらしい。
角状突起の位置にも種類による違いがある。アリオラムス・アルタイの角状突起は、腹方突起の背側(真上)に位置している。これはタルボサウルス、ティラノサウルス、大部分のダスプレトサウルスの成体、テラトフォネウスなどと同様である。それに対してアルバートサウルス、アパラチオサウルス、ビスタヒエヴェルソル、ゴルゴサウルス亜成体、ダスプレトサウルス幼体などでは、角状突起の頂点が腹方突起よりも前方にある。
アリオラムス・アルタイでは角状突起の頂点が1つで、真上を向いている。これはテラトフォネウスと同じである。一方、アルバートサウルス、ゴルゴサウルス、ダスプレトサウルスの亜成体では、頂点が2つか3つあり前背方を向いている。

ゴルゴサウルス亜成体

ダスプレトサウルス亜成体?

テラトフォネウス幼体
コメント ( 2 ) | Trackback ( 0 )