獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年9月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ディウキン

Diuqin は現地のマプチェ語で猛禽の意であるという。このqiをなんと読むか問題は、中国語のピンインならチであるが、マプチェ語でどう発音するのかわからない。英語の動画ではダイウクィンなどと言っていたので、仮にディウキンとしておく。
ディウキンは、後期白亜紀サントニアン(Bajo de la Carpa Formation)にアルゼンチンのネウケン州に生息した中間的なウネンラギア類で、2024年に記載された。サントニアンの地層から初めて見つかったウネンラギア類であり、形態学的にもチューロニアンーコニアシアンのウネンラギアなどとカンパニアンーマーストリヒティアンのアウストロラプトルの中間的な形質を示す。ウネンラギア類の進化史においてギャップを埋めるものと考えられている。
ディウキンのホロタイプ標本は断片的な胴体の骨格で、最後の仙椎の神経弓、前方の尾椎の神経弓、ほとんど完全な上腕骨、いくつかの未同定の骨からなる。どうしてこれだけでウネンラギア類とわかるのだろうか。
ディウキンの上腕骨は近位端以外はほぼ完全で、上腕骨全体に対する三角筋稜deltopectoral crestの短さ、三角筋稜にdistolateral deltopectoral ridge という稜があることなど4つの特徴から、ウネンラギア類と同定された。これらの特徴はブイトレラプトルやウネンラギアにもみられる。
ディウキンの上腕骨はブイトレラプトルよりは太く、アウストロラプトルよりは細く、太さはウネンラギア・コマフエンシスと最も近い。シグモイド形(S字状)のカーブは、ウネンラギア類の中ではアウストロラプトルと似ている。ディウキンの三角筋稜が前方を向いていることは、アウストロラプトルと似ており、三角筋稜が前側方を向いているブイトレラプトルやウネンラギアとは異なる。このようにいくつかの点でアウストロラプトルと似ていることから、ディウキンはウネンラギアなどとアウストロラプトルの中間の形質を示すと考えられた。ディウキンの推定全長はウネンラギアと同じくらいで、アウストロラプトルの祖先形に近いものかもしれない。
ディウキンの上腕骨の遠位端には、2つの円形に近い孔があり、咬み跡と考えられる。ブイトレラプトルのホロタイプ標本にも同じような孔があり、ワニ類か哺乳類によるものと考えられている。骨が治癒した跡はみられないことから、死後あるいは死の直前の咬み跡と思われる。
ディウキンのホロタイプ標本から2-3 mの位置から、分離した歯が見つかっている。この歯は保存された部分が22.69 mmで、歯冠の先端1/3が欠けている。歯冠はほどほどに側偏し、前縁が凸型で後縁が凹型にカーブしている。後縁が凹型にカーブしていることは、オルコラプトル、メガラプトル、ムルスラプトルのようなメガラプトル類と同様であり、後縁がまっすぐなアベリサウルス類とは異なる。また、前縁には鋸歯がなく、後縁にのみ鋸歯があることは、派生的なメガラプトル類の特徴である。またウネンラギア類を含むパラヴェス類の特徴は示していない。これらのことから、この歯はメガラプトル類と推定される。同じBajo de la Carpa Formationからはメガラプトル類トラタイェニアの化石が見つかっていることから、トラタイェニアの可能性が高いと著者らは考えている。
参考文献
Juan D. Porfiri, Mattia A. Baiano, Domenica D. dos Santos, Federico A. Gianechini, Michael Pittman and Matthew C. Lamanna. (2024) Diuqin lechiguanae gen. et sp. nov., a new unenlagiine (Theropoda: Paraves) from the Bajo de la Carpa Formation (Neuquén Group, Upper Cretaceous) of Neuquén Province, Patagonia, Argentina. BMC Ecology and Evolution (2024) 24:77
コメント ( 0 ) | Trackback ( 0 )
シュリ・デヴィ
Copyright 2023 Czepiński
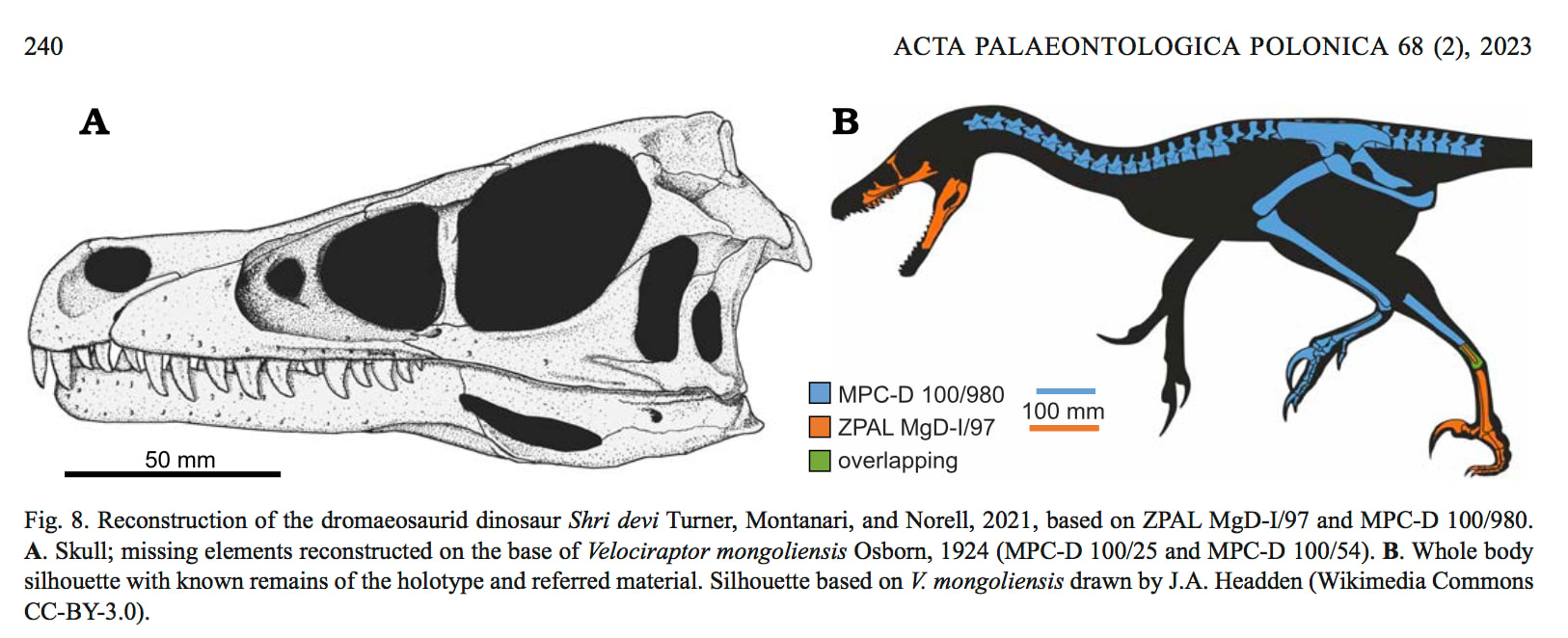
シュリ・デヴィは、後期白亜紀カンパニアン(バルンゴヨット層Baruungoyot strata)にモンゴルのゴビ砂漠のフルサンKhulsanに生息したヴェロキラプトル亜科のドロマエオサウルス類で、2021年に胴体の部分骨格が記載されたが、頭骨は発見されていなかった。その後、関節した足と部分的な頭骨が発見され、2023年に記載された。
モンゴルのゴビ砂漠からは、ヴェロキラプトルに似たドロマエオサウルス類の化石が多数発見されてきた。ヴェロキラプトル・モンゴリエンシス、ヴェロキラプトル・オスモルスカエ、ツァーガン、リンへラプトルなどである。1980年代にVelociraptor sp.とされた部分骨格がTurner, Montanari and Norell (2021) により、胴椎や足の特徴にもとづいて新属新種Shri deviとして記載されたが、頭骨はなかった。このホロタイプ標本は頸椎、胴椎、前方の尾椎、右の大腿骨、左右の脛骨、右の足からなる。
Czepiński (2023) は関節した足と部分的な頭骨を発見し、足の特徴が一致することからシュリ・デヴィと同定した。この新しい標本ZPAL MgD-I/97は、左の頬骨、涙骨、上顎骨、右の上顎骨の断片、口蓋部分、左右の歯骨、夾板骨、上角骨、角骨からなる部分的な頭骨と、左の後肢(脛骨、腓骨、距骨、完全な中足骨と指骨)が密着したものである。
他のドロマエオサウルス類と異なるシュリ・デヴィの特徴は、胴体では第1胴椎のエピポフィシスが大きく、後関節突起の上にかぶさることや、後肢の第2指の末節骨(カギ爪)が相対的に大きいことなどである。また新しい標本からわかった頭骨の特徴は、前眼窩窓が短いこと(前後の長さと背腹の高さが同じ)、前眼窩窩の腹側縁に沿って神経血管孔のすぐ上にはっきりした稜があること、上顎骨と頬骨の縫合線がZ形であることなどである。
シュリ・デヴィは他のヴェロキラプトル亜科(ヴェロキラプトル、ツァーガン、リンへラプトルなど)と比べて吻が短い。前眼窩窓の長さ/高さの比率は、シュリ・デヴィでは1.0 であるがヴェロキラプトル・モンゴリエンシスでは1.3-1.5、リンへラプトルで1.5 である。これについては成長段階の違いとも疑われるが、シュリ・デヴィの頭骨全体はヴェロキラプトルなどとほとんど同じくらい大きく、成長段階の差では説明できないとしている。
この吻が短いことについては、北アメリカのアトロキラプトルなどサウロルニトレステス亜科との収斂といっている。ただしシュリ・デヴィの頭骨は少し短いとはいってもスレンダーであり、それほど頑丈なわけではない。過去の研究ではモンゴルのような砂漠の環境ではトカゲや哺乳類など小型の獲物を捕食するため吻が細長く、北米のより湿潤な環境ではもう少し大型の獲物に対応するため吻が短めという説明であった。シュリ・デヴィのバルンゴヨット層は、ヴェロキラプトル・モンゴリエンシスのジャドフタ層ほど乾燥気候ではなかったという。しかし小型や中型の脊椎動物相は大きな違いはなく、獲物による違いとはいいきれないともいっている。また鼻腔の容積は体温調節と関連することから、そのため気候の違いと関係するかもしれないという。
確かに、少し吻が短いヴェロキラプトルみたいなものであるが、ヴェロキラプトル属の別種ではいけないのだろうか。やはりリンへラプトルとツァーガンの問題も含めて、ヴェロキラプトル亜科のメンバーについては再検討が必要なのだろう。
参考文献
Czepiński, Ł. 2023. Skull of a dromaeosaurid dinosaur Shri devi from the Upper Cretaceous of the Gobi Desert suggests convergence to the North American forms. Acta Palaeontologica Polonica 68 (2): 227–243.
コメント ( 0 ) | Trackback ( 0 )
ナトヴェナトル
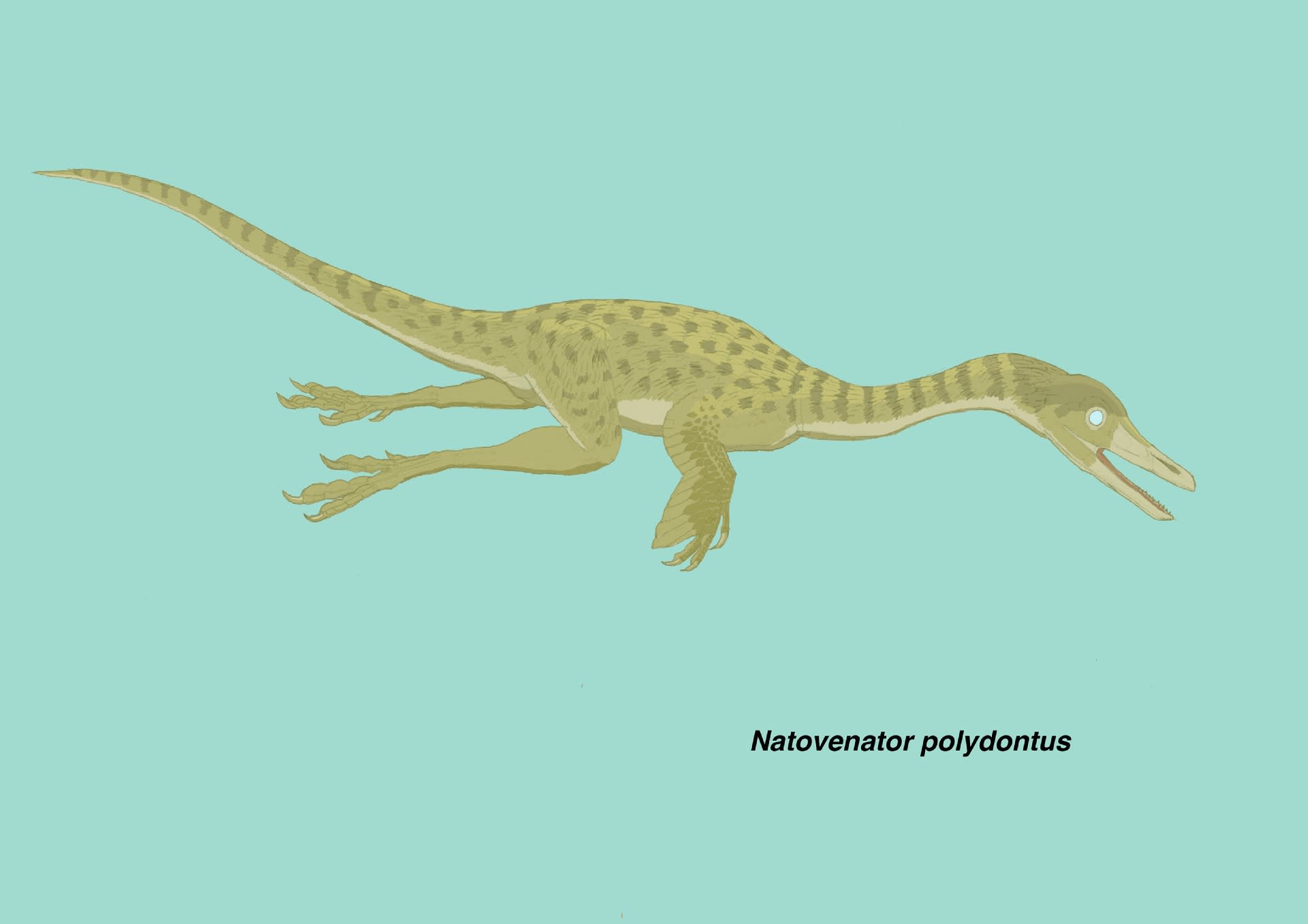
ナトヴェナトル・ポリドントゥスNatovenator polydontusは、後期白亜紀(Baruungoyot Formation)にモンゴルのゴビ砂漠(Hermiin Tsav)に生息したドロマエオサウルス類(ハルシュカラプトル亜科)で、2022年に記載された。ホロタイプ標本は、ほとんど完全な頭骨を含む、大部分が関節した骨格である。ハルシュカラプトルと似ており、同様に半水生の生活をしていたと考えられた。
この発見により改訂されたハルシュカラプトル亜科の特徴は、背腹に扁平な前上顎骨、前上顎骨に多数の神経血管孔がある、前上顎骨歯が大きく密集し、遅延した置換パターンをもつ、前方の上顎骨歯が縮小している、外鼻孔が後退し背外側にある、非常に長く伸びた頸椎、前方の頸椎の後関節突起が癒合して葉状になっている、尺骨が扁平となり鋭い後縁をもつ、第2と第3中手骨が同じ太さである、などである。
ハルシュカラプトルとも異なるナトヴェナトルの固有派生形質は、前上顎骨の前端近くに幅広い溝がある;前上顎骨の鼻孔間突起internarial processが長く、鼻骨を覆い外鼻孔の後方まで伸びている;大きく切歯形の13本の前上顎骨歯;最前方の3本の上顎骨歯が縮小し密集している;外鼻孔が前後に長い(眼窩の前方部の30%);軸椎の側面に後側方を向いた突起がある;頸椎にプレウロシールがない;肋骨の基部が後側方を向き、また水平に近い;第2中手骨の中央がくびれているhourglass-shapedなどである。
前上顎骨には多数の歯があり、歯槽が骨の隔壁で仕切られることなく密集している。歯冠はハルシュカラプトルと同様に高く切歯形である。置換歯が大きいことから、前上顎骨歯の置換はハルシュカラプトルと同様に遅いと考えられた。ただしナトヴェナトルでは前上顎骨歯は13本で、ハルシュカラプトルの11本より多い。上顎骨では、最も前方の3本の歯が、前上顎骨歯や後方の上顎骨歯よりも顕著に小さい。このパターンはハルシュカラプトルにもみられるが、ハルシュカラプトルでは縮小した歯は2本である。上顎骨と歯骨の歯は鋭い牙状で鋸歯がなく、それぞれ23本以上と考えられた。
頸椎は10個だが一個一個が長く伸びている。そのためほとんどのドロマエオサウルス類よりも首が長く、頸椎全体は胴椎全体よりも長い。しかしハルシュカラプトルよりは短い(ハルシュカラプトルでは頸椎全体が胴椎+仙椎と同じ)。頸椎にはプレウロシールがない。一方ハルシュカラプトルでは、第7から第9頸椎にプレウロシールがある。
ナトヴェナトルの胴椎の肋骨は、かなり後側方を向いている。これは地上性の獣脚類と異なり、ウミスズメ、ウミガラス、アビ、ヘビウのような多くの潜水性の鳥類と似ている。これらの潜水性の鳥類では、後方を向いた肋骨は体を流線形にすることで、遊泳を助けている。この後方を向いた肋骨は鳥類のほか、半水生のカモノハシやタニストロフェウスにもみられる。一方、モササウルスやクジラ類のような完全に水生の四肢動物では、胸椎全体が傾いたり、胸郭が前方に集中することで全体として流線形を作り出すのに役立っている。つまりナトヴェナトルの肋骨は、半水生の動物の傾向を表している。
ナトヴェナトルの肋骨のもう一つの特徴は、基部が水平に近く幅広いアーチをなすことで、これは背腹に圧縮された樽状のbarrel-shaped胸郭を表す。これも半水生と推定されるスピノサウルス類やコリストデラ類にみられるという。このようにナトヴェナトルの肋骨はいずれも半水生の生態を支持する。
論文の生体復元図(Fig.5)にはアヒルのように水面に浮いた姿と、潜水して魚を追いかける姿の両方が描かれている。やはり潜水して泳いでいる姿勢の方が良いと思ったので、イラストはそうした。このアヒルのように浮いた姿は、羽毛の性質による。ガンカモ類は防水性の羽毛で空気を含むので、このように浮いていられるが、ウやヘビウの羽毛は防水性ではないため濡れてしまい、長時間水面に浮いていることはできないという。
参考文献
Sungjin Lee, Yuong-Nam Lee, Philip J. Currie, Robin Sissons, Jin-Young Park, Su-Hwan Kim, Rinchen Barsbold & Khishigjav Tsogtbaatar. (2022) A non-avian dinosaur with a streamlined body exhibits potential adaptations for swimming.
COMMUNICATIONS BIOLOGY | (2022) 5:1185 | https://doi.org/10.1038/s42003-022-04119-9 | www.nature.com/commsbio
コメント ( 0 ) | Trackback ( 0 )
ダウルロン

ダウルロン・ワンギDaurlong wangiは、前期白亜紀(Longjiang Formation)に中国内モンゴル自治区に生息した小型のドロマエオサウルス類(全長150 cm程度)で、2022年に記載された。ダウルロンのホロタイプ標本はほとんど完全な全身骨格で、内臓の組織が保存されていたことから腸管の構造について論じられている。
他のドロマエオサウルス類と区別されるダウルロンの特徴は、1)前上顎骨の鼻孔下突起が細長く、外鼻孔よりもずっと後方まで延びている(これはヴェロキラプトルなどにもみられる)、2)前眼窩窩の前端に大きく台形のpromaxillary recessがある、3)maxillary fossaが大きく浅く、後方にあるのでpila promaxillarisがpila interfenestralisよりも幅広い。などである。
2)3)は用語が難解のようだが特に難しいことではない。エウドロマエオサウリアの上顎骨の図を参照していただきたい。promaxillary fenestra の奥にある空所がpromaxillary recessであるが、本文ではpromaxillary fenestraが台形と書いてあり、区別していないようにみえる。一方、maxillary fenestra を囲む浅い窪みがmaxillary fossaであるが、ダウルロンではmaxillary fenestraがあるとは書かれていない。貫通した孔はないのかもしれない。
ダウルロンのホロタイプ標本は、ティアンユーラプトルの85%、ゼニュアンロンの93%の大きさである。頭骨は大腿骨の94%の長さであり、これは多くのドロマエオサウルス類と同様である(ハルシュカラプトル、ミクロラプトル、サウロルニトレステスで92–95%)。ゼニュアンロンでは86%、ティアンユーラプトルでは頭骨が大腿骨よりも長い。吻が長いヴェロキラプトルでは117 から128%である。
前上顎骨の前縁は腹側縁と直角をなしている。これはヴェロキラプトル、ゼニュアンロン、サウロルニトレステスと同様で、鋭角をなすミクロラプトルやシノルニトサウルスとは異なる。前上顎骨の鼻孔下突起は外鼻孔の後端よりもずっと後方に延びており、これはヴェロキラプトルやリンへラプトルと同様で、ミクロラプトルやゼニュアンロンと異なる。
ゼニュアンロンと同様に、前眼窩窩は大きく上顎骨の2/3以上をカバーしている。promaxillary fenestraは比較的大きく、前眼窩窩の前腹方端に接している。シノルニトサウルスとゼニュアンロンではpromaxillary fenestraはもっと後方にある。promaxillary fenestraは台形で、まっすぐな前縁、後縁、腹側縁と後背方に傾いた背側縁に囲まれている。pila promaxillarisがpila interfenestralisよりも幅広いことは、ティアンユーラプトルともゼニュアンロンとも異なっている。ミクロラプトル類ではpila promaxillarisの周辺などに小さい孔や稜があるが、ダウルロンにはない。またゼニュアンロンでは前眼窩窩の後腹側部分に孔や稜があるが、ダウルロンにはない。
眼窩は卵形でその長軸は前腹方-後背方を向いている。強膜輪が比較的よく保存されている。歯骨は長さ/ 高さの比率が約8で、細長いミクロラプトルやヴェロキラプトルと、太いサウロルニトレステスの中間である。
前上顎骨の歯は1個だけ保存されており、鋸歯がなく先端が尖っていて、先端近くが後方に曲がっている。上顎骨には10個の歯が保存されており、あと1個が失われている。その歯冠はティアンユーラプトルよりも太く長い。中央の上顎骨歯が最も長い。一部の歯冠では後縁に鋸歯が保存されている。すべての上顎骨歯はナイフ形で、先端近くがわずかに後方に反っている。ゼニュアンロンでは後縁がもっとカーブしている。歯骨歯は6個が観察され、それらは上顎骨歯よりもずっと小さい。
前肢の長さは後肢の60%より小さい。ドロマエオサウルス類の中で比較的短い前肢は、アウストロラプトル、ハルシュカラプトル亜科、ティアンユーラプトル、ゼニュアンロンと共通している。三角筋稜は顕著に発達していない。尺骨はわずかに後方に反っており、ティアンユーラプトルよりも太い。
ダウルロンでは大腿骨の長さは脛骨の91%である。ティアンユーラプトルとゼニュアンロンでは、大腿骨は脛骨の77%以下なので、それらの方がひざ下が長いことになる。中足骨IIIとIVはほとんど同じ長さで、大腿骨の61%である。
羽毛の痕跡は、後頭部から背中にかけてと尾椎に沿って観察された。四肢の近くや体の腹側には保存されていないので、ゼニュアンロンのように翼に風切羽があったかどうかははっきりしない。背中のあたりの羽毛はシノルニトサウルスのように根元から枝分かれした形をしていたらしい。尾椎の尾椎鞘caudothecaの近くには斜め後方を向いた尾羽が観察された。
系統解析の結果、ダウルロンは、ティアンユーラプトルとゼニュアンロンと最も近縁となり、これら3種はミクロラプトル亜科とエウドロマエオサウリアの中間段階のクレードにまとめられた。これらは上顎骨の前眼窩窩が非常に拡大していること、上腕骨の三角筋稜が縮小していること、腸骨の前寛骨臼突起が長いこと、を共有している。これらの「ティアンユーラプトル様のドロマエオサウルス類」は、体のサイズがミクロラプトル亜科とエウドロマエオサウリアの中間である。またミクロラプトル亜科と共有する形質、エウドロマエオサウリアと共有する形質、さらにより基盤的なドロマエオサウルス類(ハルシュカラプトル亜科、ウネンラギア亜科)と共有する形質を、モザイク的に示している。
ダウルロンの腹腔内には、青黒い組織の遺残が保存されており、腹側はガストラリア、後方は恥骨に囲まれている。これは腸管の痕跡と考えられる。この組織の前端は第9胴椎の位置にあり、そこから第10と第11胴椎の間で最大の高さに達し、ガストラリアと接している。後方の恥骨の前縁との間には何もない隙間がある。このような配置は、スキピオニクスと同様である。ちなみにシノサウロプテリクスの1つの標本の腹腔内にある円形の構造物は、卵ではないかと言われたことがあるが、それはスキピオニクスの十二指腸とよく似ており、ダウルロンにもあることから、腸管と考えられる。基盤的なコエルロサウルス類またはアロサウロイドであるスキピオニクスと、鳥類に近いダウルロンとで腸管の分布が同様であることから、肉食の獣脚類の消化管の配置は保守的であったと考えられる。アヴィアラエになってから、鳥類特有の配置を獲得したらしいという。
ドロマエオサウルス類の中では前肢が短いグループとなったが、頭と比べると前肢が短いようにはみえない。上腕、前腕は短いが手は結構長いようにみえる。前半身が小さく後半身が大きいので、後肢が長いわりに前肢は短いという感じである。顔はヴェロキラプトルなどとは全然違って短く、シノルニトサウルスのような感じだろうか。
参考文献
Xuri Wang, Andrea Cau, Bin Guo, Feimin Ma, Gele Qing & Yichuan Liu. Intestinal preservation in a birdlike dinosaur supports conservatism in digestive canal evolution among theropods. Scientific Reports (2022) 12:19965.
コメント ( 0 ) | Trackback ( 0 )
サウロルニトレステス

サウロルニトレステスは、後期白亜紀カンパニアン(Dinosaur Park Formation)に北アメリカ(アルバータ、モンタナ)に生息した中型のドロマエオサウルス類で、Sues (1978) によって記載された。ホロタイプは30弱の骨からなる部分骨格で、その後も4回ほど部分骨格が見つかっていたが、頭骨についてはホロタイプの前頭骨や外翼状骨以外にはほとんど情報が得られなかった。2014年にアルバータ州のDinosaur Provincial Park で、ホロタイプの発見場所から1 km以内の地点から、頭骨を含む関節状態のほとんど完全な全身骨格化石が発見された。まずこの頭骨についてCurrie and Evans (2019) で記載されている。
新しく発見されたUALVP 55700は、尾椎の後端といくつかの前肢、後肢の指骨を除いて、ほとんど完全な全身骨格である。頭胴長は91 cm で、尾を含めた全長は2 m弱と推定されている。この体格はヴェロキラプトルとほぼ同じであるが、頭骨の顔の部分はヴェロキラプトルよりも短く、丈が高く、幅が広い。またヴェロキラプトルは後肢の長さが15%短いという。
他のドロマエオサウルス類と異なるサウロルニトレステスの特徴は、比較的短い頭骨(頭骨/大腿骨の比が1.0より小さい);maxillary fenestra の位置が前眼窩窩の背側縁に近い;鼻骨と涙骨に前眼窩窩と通じる内腔がある;2番目の前上顎骨歯が大きく、舌側面が扁平で、唇側と舌側の両方に縦の稜や溝がある、などである。
外鼻孔の後端は、歯のある腹側縁における前上顎骨と上顎骨の縫合線よりわずかに後方にある。これはアトロキラプトルと同様である。バンビラプトル、デイノニクス、リンへラプトル、ツァーガン、ヴェロキラプトルのような他のドロマエオサウルス類では、外鼻孔の後端はもっと後方にある。
サウロルニトレステスでは、前眼窩窩の前端が上顎骨の4番目の歯槽の上にある。バンビラプトルでは3番目の歯槽の上にある。アケロラプトルとツァーガンでは5番目の歯槽の上にあり、ヴェロキラプトルでは4番目と5番目の間にある。
前眼窩窓は大体三角形で、長さよりも高さが大きい。一方、リンへラプトル、ツァーガン、ヴェロキラプトル、デイノニクス、ドロマエオサウルスでは高さよりも長さが大きい。バンビラプトルとシノルニトサウルスでは、ほぼ同じか高さが大きい。
サウロルニトレステスの特徴としてmaxillary fenestraの位置が背側にあることを挙げているが、これを表現するのに、maxillary fenestraの腹側縁から上顎骨の腹側縁までの距離/ 最も大きい歯冠の高さ、という比率を用いている。この数値はサウロルニトレステスでは2.33 で、アトロキラプトルの2.2と近いが、ほとんどのドロマエオサウルス類よりも大きいという。この数値はバンビラプトル、デイノニクス、ドロマエオサウルス、リンへラプトル、ツァーガンでは1.5–1.8である。ヴェロキラプトルでは1.5 以下という。
眼窩の輪郭は左右両側ともゆがんでいるが、おそらく円形に近く、長さと高さがほぼ等しいと思われる。下側頭窓は縦に長く、いくらかくびれている。
右の鼻骨の後方部分は外れて背側に回転しているため、腹側面が一部見えている。この鼻骨の後端の腹側面には、3つの含気孔があって内部に通じている。これは他のどのドロマエオサウルス類でも報告されていない。ヴェロキラプトル、ツァーガン、バンビラプトルではこの部分が保存されているが、鼻骨は非常に薄く含気孔があるようにはみえない。デイノニクスの鼻骨には含気孔があるが、背外側面にあるので別のものである。
また前眼窩窩の後背方部分をなす涙骨の前方突起にも、いくつかの含気孔があり涙骨の内部に通じている。
前上顎骨に4個、上顎骨に11~12個、歯骨に15個の歯槽がある。前上顎骨の4本の歯のうち1番目は最も小さく、2番目が最も大きく、3番、4番と小さくなる。2番目の前上顎骨歯は歯冠の高さ、歯槽の前後長とも1番目の1.5倍あり、舌側面が扁平になっている。舌側面には3本の顕著な稜があり、それらは前半部に偏っている。唇側面にはかすかな5本の稜がある。稜と稜の間には縦溝fluteがある。この特徴的な形態は、過去にアルバータで発見されていたZapsalis abradens という歯化石と一致することから、Zapsalis はサウロルニトレステスの2番目の前上顎骨歯であることがわかった。
前頭骨は、サウロルニトレステスでは比較的よく見つかっている骨である。前頭骨の前端の鼻骨との関節面には複雑に入り組んだinterdigitating多数の突起がある。眼窩の間の広く平坦な領域と、上側頭窩supratemporal fossa の間には、強いS字状のカーブがある。これはサウロルニトレステス亜科とヴェロキラプトル亜科にみられる特徴で、ドロマエオサウルスではもっとかすかでS字状ではない。またこのカーブに沿って後眼窩骨との関節面の内側に、小さな窪みdepression がある。これもサウロルニトレステス亜科とヴェロキラプトル亜科にみられるもので、ドロマエオサウルスにはない。
参考文献
Currie, P. J., and D. C. Evans. 2019. Cranial anatomy of new specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta. The Anatomical Record 04715:1–25.(オンラインが2019, 紙が2020)Anat Rec, 303:691–715, 2020.
コメント ( 0 ) | Trackback ( 0 )
エウドロマエオサウリアの系統進化の新しい仮説 (3)


アルバータの研究者の興味の中心は、サウロルニトレステスの全身化石で盛り上がっているところの、サウロルニトレステス亜科にあるだろう。そこでサウロルニトレステス亜科とはどんなものか、その一端でも垣間見たい。細かい骨学的特徴は、専門の研究者にしかわからないのだと、思考停止してしまうのはあまりにもったいないことである。今回、一番面白かったのは、上顎骨だけでも系統関係の大体の傾向が読み取れることである。
上顎骨の周辺の構造をみると、まず前眼窩窓antorbital fenestra がある。前眼窩窓の周りには、前眼窩窩antorbital fossa がある。前眼窩窓は貫通した穴をさすが、前眼窩窩は浅くくぼんだ骨の部分をさし、涙骨や頰骨にもある。
上顎骨の前眼窩窩には、2つの穴がある。後方にあるmaxillary fenestra と前方にあるpromaxillary fenestra である。promaxillary fenestra は前眼窩窩の前縁に隠れていて側面からは見えないこともある。
エウドロマエオサウリアでは、maxillary fenestraが、前眼窩窩の中でもさらに浅くくぼんだ部分の中にある。この副次的なくぼみを、maxillary fossa という。ここではこれに注目する。
ヴェロキラプトル亜科では、maxillary fossa が広く後方に向かって開いている(赤い矢印)。ツァーガンではmaxillary fenestraが前方に移動しているが、 maxillary fossaの方向はヴェロキラプトルと同じである。
モンゴル産のドロマエオサウルス類でもアキロバトルでは、 maxillary fossaが後背方を向いている。北米のデイノニクスでも後背方に向かって開いている。ヴェロキラプトルなどと比べると狭くなっているが、閉じてはいない。
サウロルニトレステスやバンビラプトルでは、この maxillary fossaが後背方で閉じて、新しい含気性のくぼみを形成する(青い矢頭)。そのためmaxillary fenestraの中に2つ穴があるように見える。後背方の新しい穴は、maxillary fenestraとは独立したくぼみである。アケロラプトルとアトロキラプトルでもそうなっている。
このmaxillary fossa の状態は、各クレードの共有派生形質に含まれている。広く後方に開いたmaxillary fossaはヴェロキラプトル亜科の共有派生形質であり、閉じて含気性のくぼみを形成することはサウロルニトレステス亜科の共有派生形質である。

うちのバンビラプトルでも確かに確認できる。
コメント ( 0 ) | Trackback ( 0 )
エウドロマエオサウリアの系統進化の新しい仮説 (2)
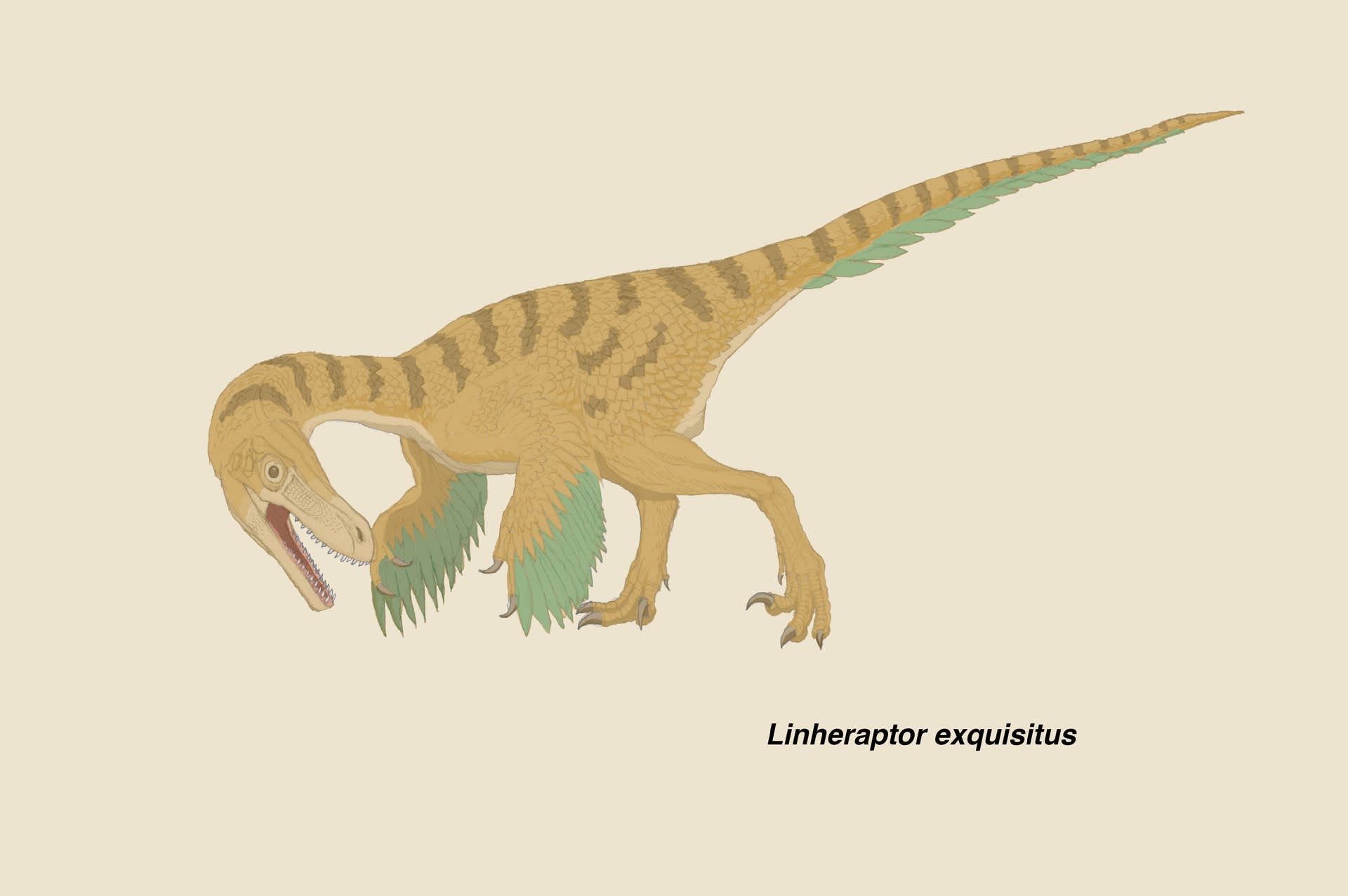
リンへラプトルは、ヴェロキラプトルと同等以上に美しい全身骨格化石である。
リンへラプトルとツァーガンの問題については、過去の記事に取り上げた。Turner et al.(2012) によって、リンへラプトルはツァーガンのシノニムであるとされたところまでである。その後、Xu et al. (2015) はさらに詳細に観察して反論している。
Xu et al. (2015) は、リンへラプトルとツァーガンの間で61の異なる形質と22の共有する形質をあげている。特に61個のうち1-8は、他のドロマエオサウルス類にはみられないリンへラプトルの固有形質であり、リンへラプトルはツァーガンとは区別できる有効な属・種であると結論している。
リンへラプトルではMaxillary fenestraが大きく、外鼻孔と同じくらいの長さである。ツァーガンや他のドロマエオサウルス類では外鼻孔より小さい。
リンへラプトルでは鼻骨の前上顎骨突起が長く伸びて外鼻孔の前端に達している。ツァーガンや他の獣脚類ではもっと短く、外鼻孔の中央付近で終わっている。
リンへラプトルでは涙骨の背側面に、いくつかの前方に開いた孔がある。これも他のドロマエオサウルス類にはない特徴で、ツァーガンやヴェロキラプトルにはこの孔はない。
リンへラプトルではMaxillary fenestraの前縁が前眼窩窩の前縁と接している。ツァーガンではMaxillary fenestraは前方にあるが、前眼窩窩の前縁とは少し離れている。
リンへラプトルでは後眼窩骨の頬骨突起と前頭骨突起の間に、はっきりした凹型のカーブがある。この点でリンへラプトルはヴェロキラプトルとは似ているが、ツァーガンなどとは似ていない。
という調子で61個列挙している。上記は写真を見てわかりやすいものである。
ただ、これらの中にはあまりに細かすぎて個体変異や保存状態の違いを拾ってしまうのではないかというものもあった。また異なる形質がいくつあり、同じ形質がいくつあるといわれても、それがドロマエオサウルス類全体の中でどの程度のものなのか、他の研究者がどうみているか、がわからなかったので、これまで記事にはしなかった。
最新のPowers et al. (2022)では、リンへラプトルとツァーガンはほとんどの形質が一致しており、異なる形質は3つのみであることから、Turner et al.(2012) の見解を支持するとしている。同種と断定しているわけではないが極めて似ている標本とみている。アメリカ自然史博物館だけでなくアルバータの研究者もそのように見ているということである。(ただしそれが正しいのかどうかはわからない。)またヴェロキラプトルが多系群になっている件も含めて、ヴェロキラプトル亜科の分類については再検討の研究が進行中とある。リンへラプトルとツァーガンについては同属別種とか、折り合いをつけて平和に解決してほしいものである。
Powers et al. (2022) は、当時のモンゴルの環境が乾燥気候であったことから、ヴェロキラプトル亜科の細長い顎は、大型動物よりも豊富な小型の哺乳類やトカゲなど、小型の獲物を捕食するのに特化したものと考えている。大型の獲物に対応したパワーよりも、小型の獲物をすばやく捕らえることを重視したということである。ただし有名な格闘化石のようにプロトケラトプスを襲った可能性もあるので、骨格だけから生態を推定するのは難しいともいっている。
まだつづく
参考文献
Xu, X., M. Pittman, C. Sullivan, J. N. Choiniere, Q.-W. Tan, J.M. Clark, M. A. Norell, and S. Wang. 2015. The taxonomic status of the Late Cretaceous dromaeosaurid Linheraptor exquisitus and its implications for dromaeosaurid systematics. Vertebrata PalAsiatica 53:29–62.
コメント ( 0 ) | Trackback ( 0 )
エウドロマエオサウリアの系統進化の新しい仮説 (1)

エウドロマエオサウリアとは、ドロマエオサウルス類の中でもウネンラギア類やミクロラプトル類などを除いた、有名なヴェロキラプトルやデイノニクスなどの最も“ラプトルらしい”肉食恐竜のグループである。中には全身骨格が発見されている種もいくつかあるが、多くは部分骨格か断片的であり、アトロキラプトルのように上顎と下顎しかないものもある。上顎骨といくつかの骨しかない種類も多いので、広範なドロマエオサウルス類の系統解析をするためには、上顎骨はかなり重要である。
Powers et al. (2022) は上顎骨に注目した。アケロラプトルの上顎骨は多くのひびが入ってつぶれており、位置関係がゆがんでいる。アトロキラプトルの上顎骨は最初の記載の時には完全にクリーニングされておらず、記載が不完全なところがあった。さらにデイノニクスの上顎骨は他のいくつかの骨とともにつぶれていたため、関節面などの構造がわからない部分があった。そこでPowers et al. (2022)はアケロラプトル、アトロキラプトル、デイノニクスの上顎骨を中心に、多くのドロマエオサウルス類で前上顎骨、鼻骨、歯骨を含めた吻部をCTスキャンし、解析した。例えばアケロラプトルについては3次元的にゆがみを直してデータを取り直した。これにより外側面と内側面の構造の比較や、内部の腔所などの構造、関節面の状態などが新たに解明された。
ドロマエオサウルス類の系統について包括的に解析した研究としてはCurrie and Evans (2019) などがあるが、本来は新しい種類が加わるときには形質の定義などを改訂すべきものである。そこで今回、CTスキャンによって得られた新知見を含めて、分岐分析に用いる上顎骨周辺の形質の表現が適切であるかどうかを注意深く再検討した。そして不適切な表現やスコアを修正して、あらためてドロマエオサウルス類の系統解析を行った。例えばヴェロキラプトルでは上顎骨全体が長いが、前方突起が「長い」ことと「細長い」ことが一緒にされていた。このままだと北米のドロマエオサウルス類はすべて「長くない」で括られてしまう。しかしサウロルニトレステスなどは前方突起が相対的に長いが、前方突起の形自体(長さと高さの比)は細長くないという。
著者らはいくつかの方法で系統解析を行っているが、いずれもエウドロマエオサウリアの中には3つのクレードが形成された。ヴェロキラプトル亜科、ドロマエオサウルス亜科、サウロルニトレステス亜科である。これらのどれが先に分岐したかは場合によって異なっていた。また、デイノニクスの位置は今回も確定せず、ドロマエオサウルス亜科の最も基盤的な位置にくる場合と、サウロルニトレステス亜科の最も基盤的な位置にくる場合があった。
ヴェロキラプトル亜科はベイズ解析の分岐図で12の共有派生形質で支持された。上顎骨に関する共有派生形質としては、広く後方に開いたmaxillary fossa、長い前方突起、細長い前方突起、広い板状のpila interfenestralis、細長い上顎骨などがある。アダサウルスでは吻部の形質はわからないが、方形頬骨や方形骨にリンへラプトルやツァーガンと共通する形質がいくつかある。最尤解析の分岐図でヴェロキラプトル・モンゴリエンシスとヴェロキラプトル sp.はクレードとなったが、ヴェロキラプトル・オスモルスカエはリンへラプトル+ツァーガンとクレードをなした。以前にも指摘されたように、これはヴェロキラプトル属が多系群であることを示唆する。
ドロマエオサウルス亜科にデイノニクスが含まれる場合は確実性は低いが、前上顎骨の外鼻孔の下の丈が高いなど、3つの共有派生形質で支持された。デイノニクスを除いたドロマエオサウルス亜科は、前縁と後縁の鋸歯がほぼ同じ大きさである、閉鎖孔突起が近位にあるなど、4つの共有派生形質で支持された。
最尤解析ではサウロルニトレステス亜科にデイノニクスが含まれたが、その場合は後縁の鋸歯が非対称な形であるなど、4つの共有派生形質で支持された。デイノニクスを除いたサウロルニトレステス亜科は確実性が高く、11の共有派生形質で支持された。サウロルニトレステス亜科の中で、アケロラプトルはアトロキラプトルと姉妹群をなした。このクレードは上顎骨の前眼窩窩が短い、前眼窩窩が歯槽の上限よりも腹側に伸びていない、という2つの共有派生形質で支持された。アケロラプトルは従来の系統解析ではヴェロキラプトル亜科に含まれていたので、この位置づけは新しいものである。
つづく
参考文献
Mark J. Powers, Matteo Fabbri, Michael R. Doschak, Bhart-Anjan S. Bhullar, David C. Evans, Mark A. Norell & Philip J. Currie (2022): A new hypothesis of eudromaeosaurian evolution: CT scans assist in testing and constructing morphological characters, Journal of Vertebrate Paleontology, DOI: 10.1080/02724634.2021.2010087
コメント ( 0 ) | Trackback ( 0 )
ユルゴヴキアとドロマエオサウルス類の尾椎の進化
Copyright 2012 Senter et al.
最新ユタラプトルの骨格に驚いてはいけないという話である。恐竜ファンなら思うだろう。ユタラプトルといえば巨大なデイノニクスのようなイメージだけど、尾椎はこれでいいのかと。
恐竜くん情報によると尾椎はそれなりに見つかっているが、今のところドロマエオ型ではないらしいということであった。BYUの最新情報は知らないので何ともいえないが、参考になる情報はある。
結論からいうと、ユタラプトルはもともと、デイノニクスのような尾ではないようだ。ではドロマエオサウルス類の中で原始的な系統かというとそうではなく、二次的に突起が短縮したと考えられている。そのことについては、聞いたこともないようなドロマエオサウルス類ユルゴヴキアの論文で研究されている。
ブイトレラプトルの尾椎は突起が短いことは知っていたが、ドロマエオサウルス類全体でどうなっているのか知らなかったので、調べてみた。
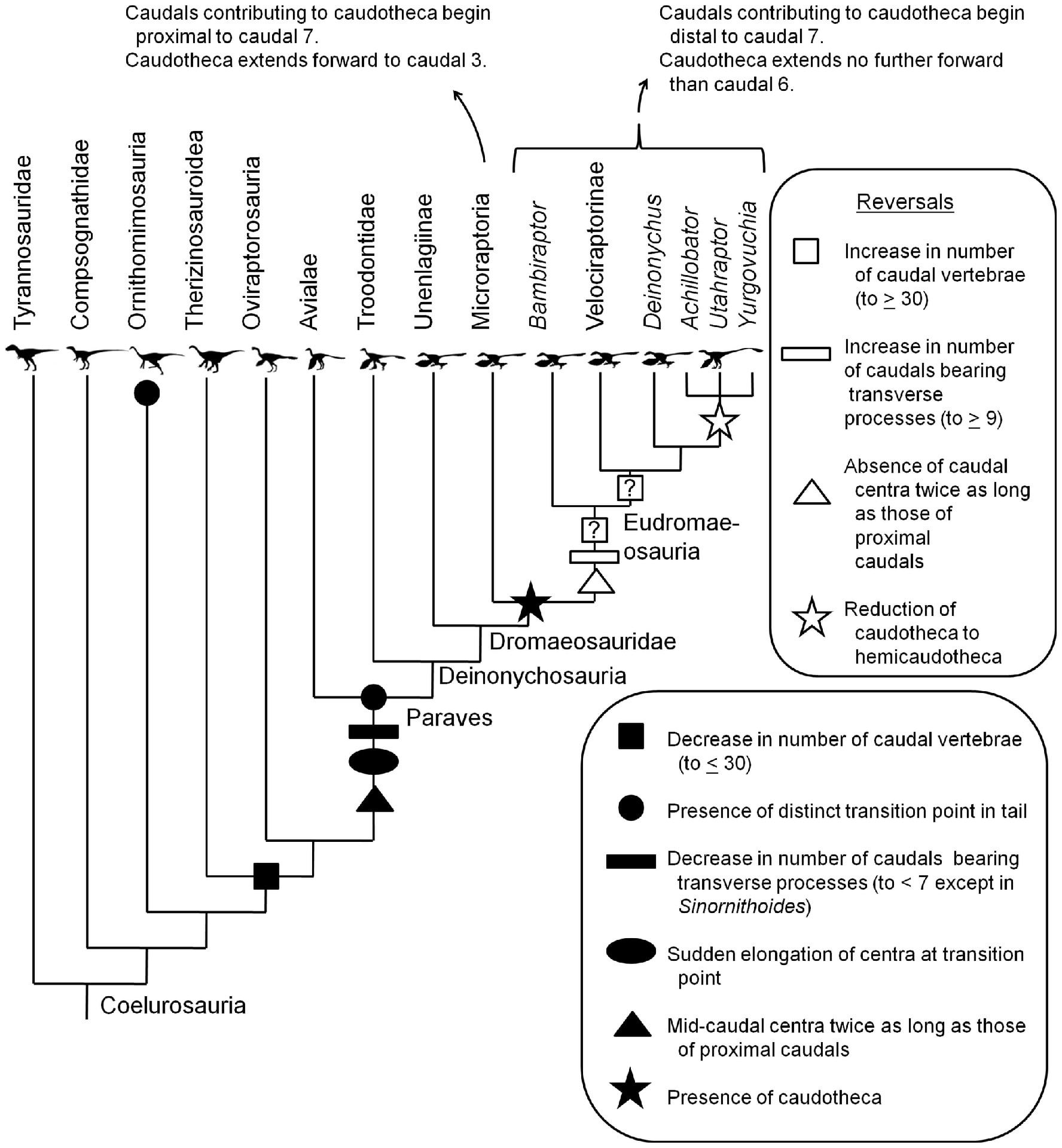
皆さんご存知のように、多くのドロマエオサウルス類では、尾椎の前関節突起と血道弓の前方突起が非常に細長く伸びて、互いに重なって尾椎を包んでいる。1個の尾椎の前関節突起は、尾椎数個分の長さに伸びている。このように尾椎を取り囲む細長い突起の束を、caudotheca (尾椎鞘)という。
しかしドロマエオサウルス類の中にも例外があり、原始的なウネンラギア類にはこの構造が発達していない。また大型ドロマエオサウルス類のユタラプトルとアキロバトルでは、中間的な形態がみられる。これらの尾椎では前関節突起が伸びているが、その長さは尾椎1個分を大きく超えることはない。つまり多少は重なるとしても、デイノニクスやヴェロキラプトルのような突起でがんじがらめの外観ではないわけである。このような状態をhemicaudotheca (半尾椎鞘)という。
Senter et al. (2012) は多くのコエルロサウルス類の尾椎を研究して、まとめた尾椎の進化シナリオを描いている。この図には多くの情報が書き込まれているが、まずは星印だけを見ていただきたい。
caudothecaはドロマエオサウルス科の中でウネンラギア類にはみられず、ミクロラプトル類以上にみられる(黒星印)。Eudromaeosauria はおなじみのヴェロキラプトル、デイノニクス、ドロマエオサウルスなどを含む進化したドロマエオサウルス類である。その中で、アキロバトル、ユタラプトル、ユルゴヴキアでは突起が短くなってhemicaudothecaになっている(白星印)。アキロバトルとユタラプトルは最も大型のドロマエオサウルス類であり、地上で方向転換などする際には尾椎がより柔軟に曲がる方が適しているのではないかと考察されている。つまり大型化にともなって尾椎の状態が祖先のコエルロサウルス類のように逆戻りreversal したといっている。
ここでは説明しきれないが、この研究にはパラヴェス類におけるtransition pointの獲得や、ミクロラプトル類とEudromaeosauriaではtransition pointの位置が異なることなど、多くの内容が含まれているので参照されたい。ドロマエオサウルス類の進化傾向の中には、多くの逆戻りreversalが含まれていることがわかる。
ユルゴヴキア自体は、ユタラプトルと同様に白亜紀前期のユタ州に生息した小型のドロマエオサウルス類である。2 m 程度の小型の種類で、属名は現地語のコヨーテに基づく。シダーマウンテン層のYellow Cat Member の下層なので、ユタラプトルより少し古い時代である。小型だが成体なので、ユタラプトルの幼体ではない。ホロタイプは頸椎、胴椎、尾椎と恥骨の一部からなる断片的なものである。尾椎のうち6個は連続しており、その中にtransition pointが含まれているので、その後方の尾椎が保存されている。前関節突起が伸びているが、中間的な状態ということである。
参考文献
Senter P, Kirkland JI, DeBlieux DD, Madsen S, Toth N (2012) New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail. PLoS ONE 7(5): e36790. doi:10.1371/journal.pone.0036790
コメント ( 0 ) | Trackback ( 0 )
ボレオニクス
大きい画像
ボレオニクスは、白亜紀後期カンパニアン後期(Wapiti Formation, Unit 3)にカナダのアルバータ中西部に生息したドロマエオサウルス類で、2016年に記載された。
アルバータ中西部のWapiti Formationには、様々な恐竜を含む典型的な白亜紀後期の動物相が保存されている。しかし種類を同定できるような特徴的な骨要素が乏しいために、Wapiti Formationの化石の分類は難しく、科レベルより細かく分類された化石はわずかしかない。これまでにハドロサウルス類エドモントサウルス・レガリスEdmontosaurus regalis と角竜類パキリノサウルス・ラクスタイPachyrhinosaurus lakustai が同定されている。パキリノサウルス・ラクスタイはパイプストーン・クリークPipestone Creekという場所のボーンベッドから、27体分もの骨が発掘されている。このパキリノサウルスの骨に混じって、いくつかの小型獣脚類の骨が発見された。研究の結果、これらの化石は新種のドロマエオサウルス類と同定されたので、Bell & Currie (2016)によって記載された。
Boreonykus certekorumの属名は「北方の爪」で、古代ギリシアの北風の神Boreasが語源らしく、種小名は発掘調査をサポートしたCertek Heating Solutions(「サーテック暖房機器」会社名?)に対する献名ということである。
ホロタイプTMP 1989.055.0047は、右の前頭骨のみである。参照標本として前肢の末節骨II-3、後肢の末節骨II-3、後方の尾椎がある。さらに多数の分離した歯が暫定的に含まれている。末節骨などは、重複した骨がないことや大きさが一致することから、暫定的にホロタイプと同一個体と考えられるが、歯についてはおそらく別個体に由来するもので、同一種と考えることには注意が必要であるといっている。
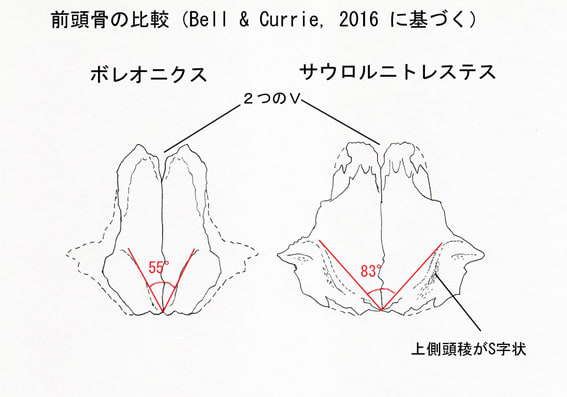
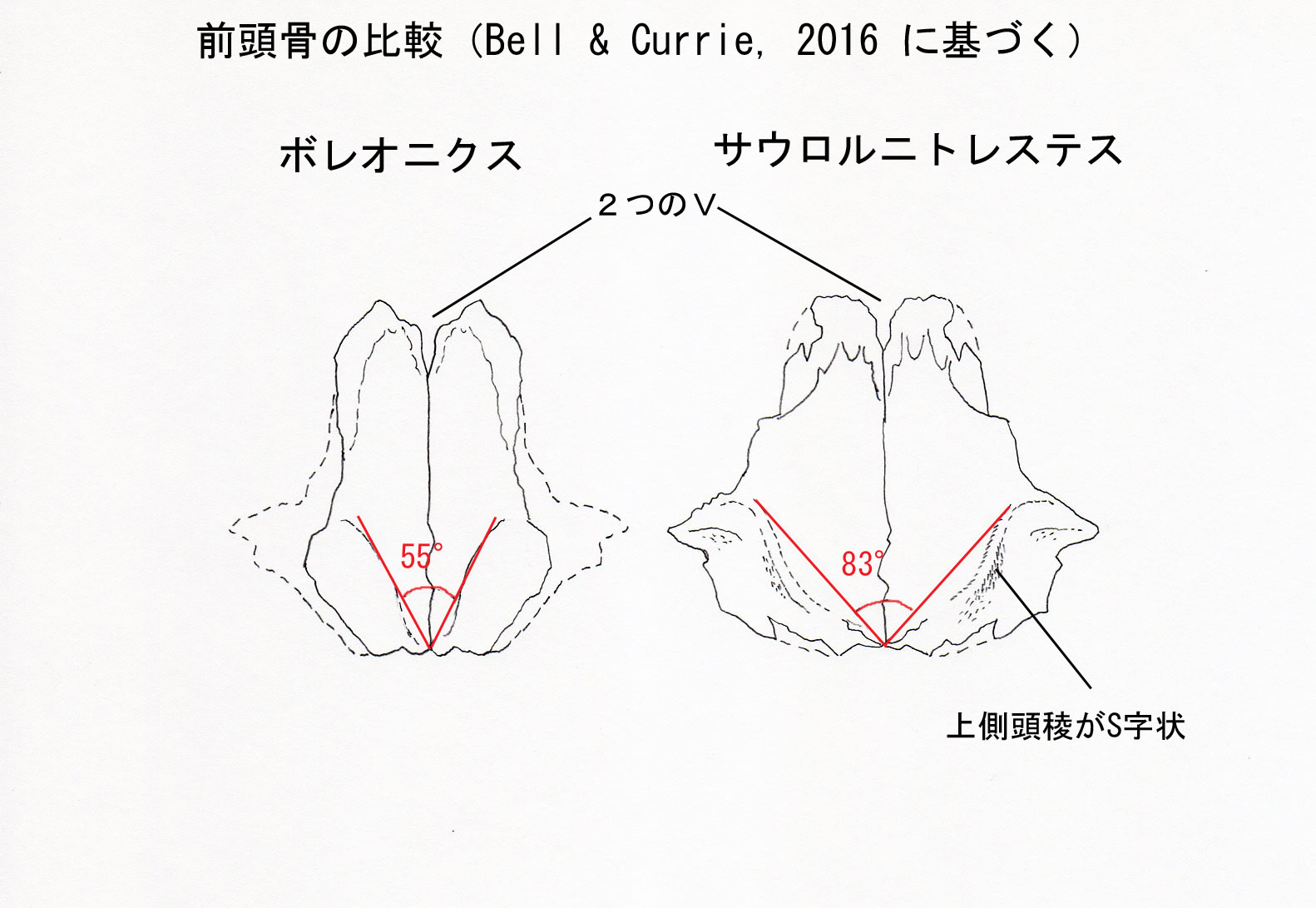
ボレオニクスの固有形質は、前頭骨の上側頭稜supratemporal ridgeが、左右を合わせると他のドロマエオサウルス類よりも鋭い角度(55°)をなすことである。また次の形質の固有の組み合わせによって他のドロマエオサウルス類と識別される。長くほっそりした前頭骨;円弧をなす低い上側頭稜;上側頭稜のすぐ後方の領域がなめらかで後腹方に傾斜している。
ボレオニクスは時代の近いサウロルニトレステスとは、よりがっしりした前頭骨、上側頭稜がS字状でないこと、前頭骨の後側方面に顕著な孔がないことで区別される。またドロマエオサウルスとは、長くほっそりした前頭骨、背面が平坦なこと、前方内側が鼻骨によって広く離れていないことで区別される。
ホロタイプは完全に近い右の前頭骨であるが、側面と後眼窩骨突起は破損している。前後に長い形状はサウロルニトレステスと似ており、比較的短いドロマエオサウルスの前頭骨とは異なっている。しかし、ボレオニクスの前頭骨は同じ大きさのサウロルニトレステスの前頭骨よりもがっしりしている(骨の厚みのようなことか)。背側からみると、ボレオニクスの前頭骨では前方の鼻骨との関節面が、2つのVが連なったような形をしている。(前方の幅広いVと後方の深いVである。)この形はサウロルニトレステスと似ている。一方、ドロマエオサウルスではこの形状が誇張されている(exaggerated とは、前方のVと後方のVがよりはっきり分離しているということらしい)。この形はツァーガンやヴェロキラプトルの鼻骨/前頭骨縫合とも異なっている。
左右の前頭骨の縫合面interfrontal sutureのうち前方2/3の部分には、長くのびた溝があり、おそらく反対側の前頭骨の稜と結合していたと思われる。このようなtongue and groove構造によって、ドロマエオサウルス類の前頭骨はトロオドン類の前頭骨と区別される。トロオドン類の前頭骨では、この縫合面が全長にわたって、細かく指状に入り組んでいるfinely interdigitated 。
後眼窩骨突起よりも前方では、前頭骨の背面はサウロルニトレステスと同様に平坦である。ドロマエオサウルスでは、ここに顕著な縦の溝sulcus がある。その結果、ボレオニクスの前頭骨はドロマエオサウルスにあるような矢状稜midline crestを形成しない。
後方には上側頭窩の前内側縁をなす上側頭稜がある。この上側頭稜は正中線に向かって凹形のカーブをなす。このカーブはドロマエオサウルスとは似ているが、サウロルニトレステスのS字状のラインとは大きく異なっている。ボレオニクスでは、上側頭稜のはさむ角度が、他のすべてのドロマエオサウルス類よりも鋭い(55°)。
参照標本の後肢の末節骨II-3は、ドロマエオサウルス類に典型的な大きく発達したカギ爪である。それは薄く、強くカーブしており、外周に沿って測ると82 mmある。内側溝medial grooveと外側溝lateral grooveは非対称であり、内側溝は外側溝よりも背側を走っている。この非対称性は派生的なドロマエオサウルス類に典型的なものであり、原始的なミクロラプトル類ではより対称に近い。また、メインの溝の腹側に、より短くはっきりしない溝がみられるが、これもドロマエオサウルス、サウロルニトレステス、ヴェロキラプトルのような派生的なドロマエオサウルス類に典型的なものである。屈筋結節はバンビラプトルよりはよく発達しているが、サウロルニトレステスやヴェロキラプトルと同様である。
ホロタイプと参照標本(分離した歯も含む)すべての標本を用いて系統解析した結果では、ボレオニクスはヴェロキラプトル亜科Velociraptorinaeとされるクレードに含まれた。ここではボレオニクスは、ヴェロキラプトル、ツァーガン、アダサウルス、アケロラプトルとポリトミーをなしている。しかし今回も含めカリーらの研究ではLongrich and Currie (2009)などのデータマトリクスに基づいているため、アメリカ自然史博物館のTurner et al. (2012) などの分岐図とは内容が全く異なっている。今回の分岐図ではヴェロキラプトル亜科とドロマエオサウルス亜科を合わせたクレードよりも外側に、アトロキラプトルとデイノニクスがきており、バンビラプトルとサウロルニトレステスはさらに外側に位置している。依然としてエウドロマエオサウリアの系統関係については、なかなか意見が一致しないようである。
また、著者自身が注意が必要と述べている、分離した歯を除いて系統解析すると、ヴェロキラプトル亜科もドロマエオサウルス亜科も崩壊してしまったという。largely similar といっているが図は示していない。エウドロマエオサウリアの一種というくらいしかわからないということらしい。やはり断片的なものは難しいのだろう。
参考文献
Phil R. Bell & Philip J. Currie (2016) A high-latitude dromaeosaurid, Boreonykus certekorum, gen. et sp. nov. (Theropoda), from the upper Campanian Wapiti Formation, west-central Alberta. Journal of Vertebrate Paleontology, 36:1, e1034359, DOI: 10.1080/02724634.2015.1034359
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ |