獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2012年5月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | ||||
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| 13 | 14 | 15 | 16 | 17 | 18 | 19 | ||
| 20 | 21 | 22 | 23 | 24 | 25 | 26 | ||
| 27 | 28 | 29 | 30 | 31 | ||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ラプトレックスは白亜紀後期のティラノサウルス科の幼体である
これはニュースとしてご存知の読者もおられると思うが、まさに表題のような論文が出ている。このサイトで以前、ラプトレックスを紹介した関係上、同様に解説を試みたい。もともと非合法にマーケットに流出し、産地と時代が推定でしかないラプトレックスは、「恐竜の成長」で盛り上がるホーナー博士のグループにとって格好の題材(標的?)になったようである。
そもそもラプトレックスの発見(Sereno et al. 2009)がScienceという一流誌に載ったのは、新種の発見だからでも保存の良い美しい骨格だからでもない。ティラノサウルス科に特徴的な形質が、白亜紀前期の小型の種類で獲得されていた、という意外性と話題性のためである。その根拠として、2つのことが確立していなくてはならない。1)ラプトレックスが産出したのが白亜紀前期の地層であること、と2)小型ではあるが、幼体でなく亜成体または成体であること、である。ホーナー博士らの論文はこの2点を論破し、完全否定している。
1)本当に白亜紀前期といえるか
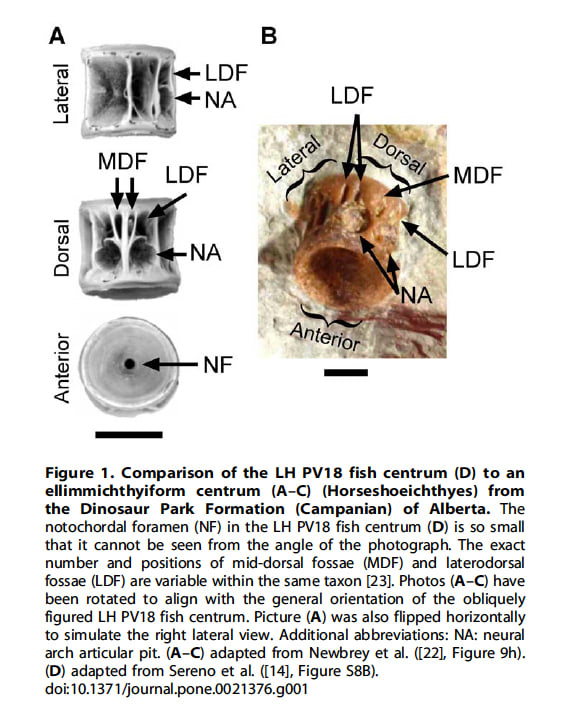
ラプトレックスの論文では、遼寧省の白亜紀前期の熱河層群・Lujiatun Beds(オーテリーヴ期からバレム期、130 Ma)から発見されたことになっている。シカゴ・トリビューンの新聞記事ではセレノ博士は「ラプトレックスの化石の母岩から回収された堆積物、魚の骨、カメ、二枚貝、その他の動物相から、遼寧省と内モンゴルの境界地域と特定できる」と述べているが、実際にサイエンスの論文で提示された証拠は、つぶれた二枚貝と魚の脊椎骨(椎体)だけである。二枚貝は同定していないので、ラプトレックスの化石の年代は、この魚の椎体に基づいていることになる。セレノらはこの魚の椎体を、白亜紀前期の硬骨魚Lycopteraと同定し、白亜紀前期の地層である根拠としている。
ところが、セレノらはLycopteraと同定した根拠を示していない。Lycoptera の脊椎骨について記述した唯一の文献はZhang (2002)であるが、これをみると問題の魚の椎体はLycopteraと非常に異なっており、Lycopteraであるはずがない、とホーナーらは述べている。ラプトレックスと共に発見された魚の椎体は、むしろEllimmichthyiformes や Hiodontiformesのものと似ている。Lycopteraの椎体は壁が薄く、円筒形で大きな脊索孔notochordal foramenをもつ点で変わっている。一方、問題の魚の椎体とEllimmichthyiformes や Hiodontiformesのものは深く両凹型で、脊索孔に対して壁が厚い。問題の魚の椎体とEllimmichthyiformes の椎体には、正中背側窩と外側背側窩 mid-dorsal and lateral-dorsal fossae があるが、Lycopteraの椎体にはこれらの凹みはない。
さらに問題の魚の椎体は、Lycopteraの椎体よりもずっと大きい。Lycopteraは小さな魚で、その椎体は典型的には1~2mm以下である。一方、問題の魚の椎体は直径4mm、長さ4mmあり、Lycopteraの椎体にしては異常に大きいが、Ellimmichthyiformes の椎体としては十分当てはまる大きさである。
問題の魚の椎体は、もっと広汎な研究をしないと正確な同定はできないが、形態と大きさからLycopteraの椎体とは考えられず、むしろEllimmichthyiformes の椎体と一致する。中国におけるEllimmichthyiformesの生息年代は、白亜紀前期から第三紀始新世にわたっているので、白亜紀前期の地層である根拠にはならない。よってラプトレックスが白亜紀前期の層から発見されたという層序学的証拠はない。
また、関係者の証言では、ラプトレックスの化石は中国ではなくモンゴルで採集されたようである。化石販売業者Hollis Butts氏によると、もともとラプトレックスの化石はモンゴルの化石発掘業者から、アメリカ人ブローカーが購入して米国に持ち込んだもので、中国由来のものと提示されたことはないという。Hollis Butts氏からKriegstein氏が買った時点では、タルボサウルスの幼体とされていたことは確認されている。どのようにして中国の白亜紀前期のものと考えられるようになったのかは、よくわからないという。
この辺の事情はどうなっているのだろうか。中国の研究機関に返還することを条件に研究が開始されたのだから、その時点でセレノ博士は中国という情報を持っていたはずではなかったか。
Copyright 2011 Fowler et al.
2)本当に成体といえるか
セレノらの論文で公表された骨学的データから、ラプトレックスの標本が成熟に近いとは結論できないという。ティラノサウルス類の個体発生において、鼻骨間の縫合は非常に早くから癒合しはじめるので、その部分的な癒合は成熟を示すものではない。
ワニの個体発生の研究から、脊椎骨の神経弓と椎体の間の縫合neurocentral sutureが恐竜の成長段階の指標として用いられる。セレノらの論文では、ラプトレックスの標本の頸椎、前方の胴椎、仙椎で神経弓/椎体縫合が「閉鎖している」ことが成熟に近いことを示唆するとされている。しかしホーナー側の研究者の一人がラプトレックスの標本を観察した結果、頸椎と前方の胴椎の神経弓/椎体縫合は、部分的に閉鎖しているだけであった。また後方の胴椎と前方の尾椎では神経弓/椎体縫合は閉じておらず、いくつかの骨は少し外れていた。さらに、神経弓/椎体の癒合はティラノサウルス科では様々な状態がみられる形質で、成熟の程度を推定するにはあまり信頼できないと考えられる。例えば、非常に大型の成体であるFMNH PR2081(スー)では、仙前椎から仙椎、尾椎にかけて閉じていない神経弓/椎体縫合がみられるという。タルボサウルスの幼体であるIVPP V4878(シャンシャノサウルス)の頸椎と胴椎では癒合の状態はさまざまで、いくつかは明らかに閉じており他のものは閉じていない。アリオラムス・アルタイの模式標本IGM 100/1844は幼体であるが、頸椎の神経弓/椎体縫合は閉じている。神経弓/椎体縫合の閉鎖はこれらのティラノサウルス科の幼体にもみられることから、ラプトレックスが成体であることを支持するものではない。
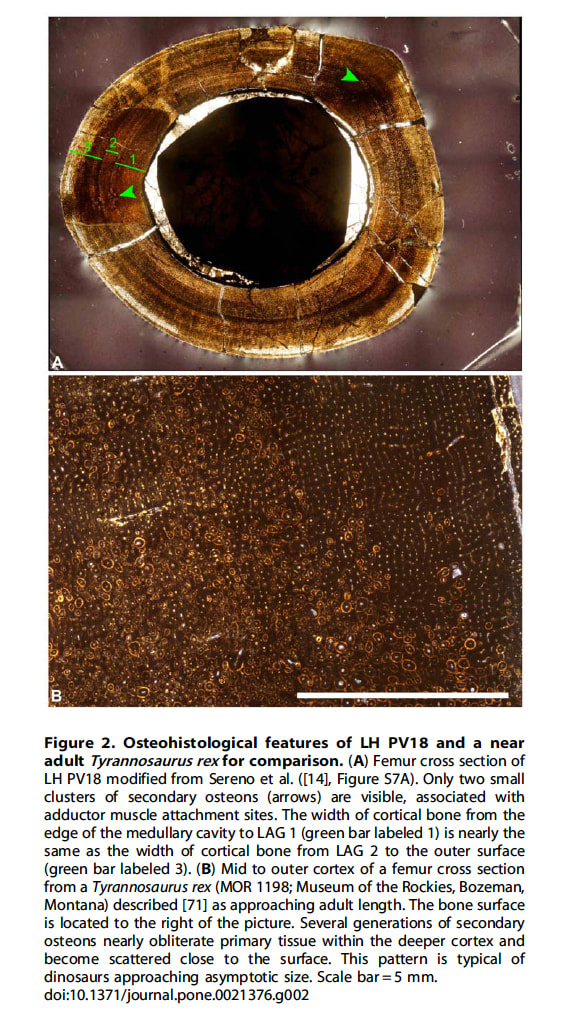
セレノらは、ラプトレックスの右大腿骨の組織切片を観察し、この個体が成熟に近い亜成体であると結論している。しかし、この切片は典型的な幼体の組織学的特徴を示している。もしこの個体が成熟に近いのならば、骨組織のリモデリングに伴う二次オステオンが、組織切片上に広範にみられるはずである。しかし二次オステオンは、2つの非常に小さい領域にしかみられない。さらに、この大腿骨は全体に叢状の繊維層板骨plexiform fibro-lamellar boneから成っているが、これは未熟な個体での急速な成長を示すものである。
長骨の断面には年輪状の成長停止線(line of arrested growth, LAG)があり、その間隔が一年分の成長量を表す(成長帯growth zone)。セレノらは、LAG1とLAG2の間に比べて、LAG2から外表面までの距離が大きくなっていることを、急激な成長期への移行と解釈しているが、これは疑問である。最も内側の骨髄腔からLAG1までの距離も、LAG2から外表面までの距離と同じくらい大きいからである。つまりLAG1とLAG2の間の成長帯は、より厚い2つの成長帯に挟まれていることになり、これは変則的な状態である。もしもLAG2から外表面までの距離を急激な成長期と考えるなら、この個体は未成熟と考えるべきである。亜成体ならば成長速度が低下している(間隔が狭まっていく)過程が観察されるはずだからである。このように、組織構築からもLAGの間隔からも、成長速度が低下している兆候は全くみられない。むしろ、外表面の骨膜の位置まで繊維層板組織が続いていることから、死亡時まで活発な成長が進行していたと考えられる。
セレノらは、この組織切片からラプトレックスの年齢を5~6才と推定しているが、これは過大評価と考えられるという。一般に年齢は、骨にみられるLAGの数を数え、それに内側からの骨吸収によって失われたLAGの数を推定した値を足して算出される。
この切片では前述のように、骨髄腔からLAG1までの間隔(図の緑の数字「1」)、LAG1とLAG2の間隔(「2」)、 LAG2から外表面までの間隔(「3」)が「厚い、薄い、厚い」となっている。セレノらは、失われたLAGの数を推定するために最も薄い「2」を用いているが、これは不適切であり、内側の「1」を用いる方が適切である。また、大型恐竜では最初のLAGが形成されるまでの間にかなり成長することを考えると、失われた部分にはLAGがなかった可能性も充分あり、その場合はラプトレックスの年齢は2~3才となる。実際、このことはラプトレックスとほぼ同じ大きさのタルボサウルスの幼体MPC-D 107/7の解析からも支持される。MPC-D 107/7の脛骨と腓骨の切片の組織学的解析から、失われたLAGはなくこの幼体は2~3才と考えられている。このように幼体に典型的な組織像とLAGの状態から、ホーナーらはラプトレックスの推定年齢として2~3才の可能性が最も高いと考えている。
Copyright 2011 Fowler et al.
3)系統分類上の問題
Tsuihiji et al. (2011) はタルボサウルスの幼体MPC-D 107/7の記載論文の中で、ラプトレックスとタルボサウルスの幼体を比較して論じている。
セレノらの系統解析では、ラプトレックスはティラノサウルス科全体の外側(基盤的な位置)にきているが、ラプトレックスが原始的であることを示す頭骨の形質10個のうち、7個はタルボサウルスの幼体MPC-D 107/7にもみられるという。このことからTsuihiji et al. (2011)は、他に何らかの基準を定めない限り、原始的な特徴と幼体の特徴を区別することは困難であると述べている。また、ラプトレックスに固有とされた形質のうち2つ(頬骨の眼窩の下部が細いこと、maxillary fenestraの背側に副次的な含気窩があること)は、MPC-D 107/7にもみられるという。そこでMPC-D 107/7をセレノらのデータに加えて系統解析にかけてみると、MPC-D 107/7はティラノサウルス科の外側、つまりラプトレックスのすぐ隣に位置した。この結果から、もしティラノサウルス科の幼体を成体と同列に系統解析すると、同じ種類の成体よりもずっと基盤的な位置にきてしまうことが確認された。よってラプトレックスがもっと派生的なティラノサウルス科の幼体である可能性があるという。
ただし、ラプトレックスとタルボサウルスの幼体MPC-D 107/7はほとんど同じ大きさであるが、両者には形態学的に異なる点がいくつかある。頭骨の細かい形質が2点と、セレノらの報告によるとラプトレックスでは腸骨の寛骨臼の背方に垂直な稜がないことである。腸骨の垂直な稜はティラノサウルス上科の共有派生形質で、重要な点である。Tsuihiji et al. (2011) は、白亜紀前期の遼寧省という時代と産地をふまえた上で、これらの形態学的な差異から、暫定的にラプトレックスはタルボサウルス・バタールの幼体ではないだろうと述べている。
これについてホーナーらの論文では、白亜紀前期の中国遼寧省という点は既に根拠が否定されていること、ホーナー側の研究者がラプトレックスの標本を実際に観察したところ、腸骨にはかすかに垂直な稜がみとめられたことを指摘している。頭骨の2つの細かい形質については、Tsuihiji et al. (2011)も同種内の変異である可能性も否定できないとしている。これらのことからホーナーらは、ラプトレックスはタルボサウルス・バタールとは限らないが、タルボサウルス属か近縁の大型ティラノサウルス類の幼体である可能性が高いとしている。
産地が不明であることと幼体であることから、ラプトレックスを完模式標本として用いることには問題がある。特に、生息年代がわからない標本は他の標本と正確に比較することができない。また幼体と成体では形態が非常に異なることはよく知られているので、ラプトレックスが本当に新種だとしても、他の種類の成体と比較することはできない。他に有力な根拠が示されない限り、ラプトレックスは疑問名nomen dubiumと考えるべきである、とホーナーらはまとめている。
参考文献
Fowler DW, Woodward HN, Freedman EA, Larson PL, Horner JR (2011) Reanalysis of ‘‘Raptorex kriegsteini’’: A Juvenile Tyrannosaurid Dinosaur from Mongolia. PLoS ONE 6(6): e21376. doi:10.1371/journal.pone.0021376
Tsuihiji T, Watabe M, Tsogtbaatar K, Tsubamoto T, Barsbold R, et al. (2011) Cranial osteology of a juvenile specimen of Tarbosaurus bataar from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. J Vert Paleontol 31: 497-517.
Copyright 2011 Fowler et al.
そもそもラプトレックスの発見(Sereno et al. 2009)がScienceという一流誌に載ったのは、新種の発見だからでも保存の良い美しい骨格だからでもない。ティラノサウルス科に特徴的な形質が、白亜紀前期の小型の種類で獲得されていた、という意外性と話題性のためである。その根拠として、2つのことが確立していなくてはならない。1)ラプトレックスが産出したのが白亜紀前期の地層であること、と2)小型ではあるが、幼体でなく亜成体または成体であること、である。ホーナー博士らの論文はこの2点を論破し、完全否定している。
1)本当に白亜紀前期といえるか
ラプトレックスの論文では、遼寧省の白亜紀前期の熱河層群・Lujiatun Beds(オーテリーヴ期からバレム期、130 Ma)から発見されたことになっている。シカゴ・トリビューンの新聞記事ではセレノ博士は「ラプトレックスの化石の母岩から回収された堆積物、魚の骨、カメ、二枚貝、その他の動物相から、遼寧省と内モンゴルの境界地域と特定できる」と述べているが、実際にサイエンスの論文で提示された証拠は、つぶれた二枚貝と魚の脊椎骨(椎体)だけである。二枚貝は同定していないので、ラプトレックスの化石の年代は、この魚の椎体に基づいていることになる。セレノらはこの魚の椎体を、白亜紀前期の硬骨魚Lycopteraと同定し、白亜紀前期の地層である根拠としている。
ところが、セレノらはLycopteraと同定した根拠を示していない。Lycoptera の脊椎骨について記述した唯一の文献はZhang (2002)であるが、これをみると問題の魚の椎体はLycopteraと非常に異なっており、Lycopteraであるはずがない、とホーナーらは述べている。ラプトレックスと共に発見された魚の椎体は、むしろEllimmichthyiformes や Hiodontiformesのものと似ている。Lycopteraの椎体は壁が薄く、円筒形で大きな脊索孔notochordal foramenをもつ点で変わっている。一方、問題の魚の椎体とEllimmichthyiformes や Hiodontiformesのものは深く両凹型で、脊索孔に対して壁が厚い。問題の魚の椎体とEllimmichthyiformes の椎体には、正中背側窩と外側背側窩 mid-dorsal and lateral-dorsal fossae があるが、Lycopteraの椎体にはこれらの凹みはない。
さらに問題の魚の椎体は、Lycopteraの椎体よりもずっと大きい。Lycopteraは小さな魚で、その椎体は典型的には1~2mm以下である。一方、問題の魚の椎体は直径4mm、長さ4mmあり、Lycopteraの椎体にしては異常に大きいが、Ellimmichthyiformes の椎体としては十分当てはまる大きさである。
問題の魚の椎体は、もっと広汎な研究をしないと正確な同定はできないが、形態と大きさからLycopteraの椎体とは考えられず、むしろEllimmichthyiformes の椎体と一致する。中国におけるEllimmichthyiformesの生息年代は、白亜紀前期から第三紀始新世にわたっているので、白亜紀前期の地層である根拠にはならない。よってラプトレックスが白亜紀前期の層から発見されたという層序学的証拠はない。
また、関係者の証言では、ラプトレックスの化石は中国ではなくモンゴルで採集されたようである。化石販売業者Hollis Butts氏によると、もともとラプトレックスの化石はモンゴルの化石発掘業者から、アメリカ人ブローカーが購入して米国に持ち込んだもので、中国由来のものと提示されたことはないという。Hollis Butts氏からKriegstein氏が買った時点では、タルボサウルスの幼体とされていたことは確認されている。どのようにして中国の白亜紀前期のものと考えられるようになったのかは、よくわからないという。
この辺の事情はどうなっているのだろうか。中国の研究機関に返還することを条件に研究が開始されたのだから、その時点でセレノ博士は中国という情報を持っていたはずではなかったか。
Copyright 2011 Fowler et al.
2)本当に成体といえるか
セレノらの論文で公表された骨学的データから、ラプトレックスの標本が成熟に近いとは結論できないという。ティラノサウルス類の個体発生において、鼻骨間の縫合は非常に早くから癒合しはじめるので、その部分的な癒合は成熟を示すものではない。
ワニの個体発生の研究から、脊椎骨の神経弓と椎体の間の縫合neurocentral sutureが恐竜の成長段階の指標として用いられる。セレノらの論文では、ラプトレックスの標本の頸椎、前方の胴椎、仙椎で神経弓/椎体縫合が「閉鎖している」ことが成熟に近いことを示唆するとされている。しかしホーナー側の研究者の一人がラプトレックスの標本を観察した結果、頸椎と前方の胴椎の神経弓/椎体縫合は、部分的に閉鎖しているだけであった。また後方の胴椎と前方の尾椎では神経弓/椎体縫合は閉じておらず、いくつかの骨は少し外れていた。さらに、神経弓/椎体の癒合はティラノサウルス科では様々な状態がみられる形質で、成熟の程度を推定するにはあまり信頼できないと考えられる。例えば、非常に大型の成体であるFMNH PR2081(スー)では、仙前椎から仙椎、尾椎にかけて閉じていない神経弓/椎体縫合がみられるという。タルボサウルスの幼体であるIVPP V4878(シャンシャノサウルス)の頸椎と胴椎では癒合の状態はさまざまで、いくつかは明らかに閉じており他のものは閉じていない。アリオラムス・アルタイの模式標本IGM 100/1844は幼体であるが、頸椎の神経弓/椎体縫合は閉じている。神経弓/椎体縫合の閉鎖はこれらのティラノサウルス科の幼体にもみられることから、ラプトレックスが成体であることを支持するものではない。
セレノらは、ラプトレックスの右大腿骨の組織切片を観察し、この個体が成熟に近い亜成体であると結論している。しかし、この切片は典型的な幼体の組織学的特徴を示している。もしこの個体が成熟に近いのならば、骨組織のリモデリングに伴う二次オステオンが、組織切片上に広範にみられるはずである。しかし二次オステオンは、2つの非常に小さい領域にしかみられない。さらに、この大腿骨は全体に叢状の繊維層板骨plexiform fibro-lamellar boneから成っているが、これは未熟な個体での急速な成長を示すものである。
長骨の断面には年輪状の成長停止線(line of arrested growth, LAG)があり、その間隔が一年分の成長量を表す(成長帯growth zone)。セレノらは、LAG1とLAG2の間に比べて、LAG2から外表面までの距離が大きくなっていることを、急激な成長期への移行と解釈しているが、これは疑問である。最も内側の骨髄腔からLAG1までの距離も、LAG2から外表面までの距離と同じくらい大きいからである。つまりLAG1とLAG2の間の成長帯は、より厚い2つの成長帯に挟まれていることになり、これは変則的な状態である。もしもLAG2から外表面までの距離を急激な成長期と考えるなら、この個体は未成熟と考えるべきである。亜成体ならば成長速度が低下している(間隔が狭まっていく)過程が観察されるはずだからである。このように、組織構築からもLAGの間隔からも、成長速度が低下している兆候は全くみられない。むしろ、外表面の骨膜の位置まで繊維層板組織が続いていることから、死亡時まで活発な成長が進行していたと考えられる。
セレノらは、この組織切片からラプトレックスの年齢を5~6才と推定しているが、これは過大評価と考えられるという。一般に年齢は、骨にみられるLAGの数を数え、それに内側からの骨吸収によって失われたLAGの数を推定した値を足して算出される。
この切片では前述のように、骨髄腔からLAG1までの間隔(図の緑の数字「1」)、LAG1とLAG2の間隔(「2」)、 LAG2から外表面までの間隔(「3」)が「厚い、薄い、厚い」となっている。セレノらは、失われたLAGの数を推定するために最も薄い「2」を用いているが、これは不適切であり、内側の「1」を用いる方が適切である。また、大型恐竜では最初のLAGが形成されるまでの間にかなり成長することを考えると、失われた部分にはLAGがなかった可能性も充分あり、その場合はラプトレックスの年齢は2~3才となる。実際、このことはラプトレックスとほぼ同じ大きさのタルボサウルスの幼体MPC-D 107/7の解析からも支持される。MPC-D 107/7の脛骨と腓骨の切片の組織学的解析から、失われたLAGはなくこの幼体は2~3才と考えられている。このように幼体に典型的な組織像とLAGの状態から、ホーナーらはラプトレックスの推定年齢として2~3才の可能性が最も高いと考えている。
Copyright 2011 Fowler et al.
3)系統分類上の問題
Tsuihiji et al. (2011) はタルボサウルスの幼体MPC-D 107/7の記載論文の中で、ラプトレックスとタルボサウルスの幼体を比較して論じている。
セレノらの系統解析では、ラプトレックスはティラノサウルス科全体の外側(基盤的な位置)にきているが、ラプトレックスが原始的であることを示す頭骨の形質10個のうち、7個はタルボサウルスの幼体MPC-D 107/7にもみられるという。このことからTsuihiji et al. (2011)は、他に何らかの基準を定めない限り、原始的な特徴と幼体の特徴を区別することは困難であると述べている。また、ラプトレックスに固有とされた形質のうち2つ(頬骨の眼窩の下部が細いこと、maxillary fenestraの背側に副次的な含気窩があること)は、MPC-D 107/7にもみられるという。そこでMPC-D 107/7をセレノらのデータに加えて系統解析にかけてみると、MPC-D 107/7はティラノサウルス科の外側、つまりラプトレックスのすぐ隣に位置した。この結果から、もしティラノサウルス科の幼体を成体と同列に系統解析すると、同じ種類の成体よりもずっと基盤的な位置にきてしまうことが確認された。よってラプトレックスがもっと派生的なティラノサウルス科の幼体である可能性があるという。
ただし、ラプトレックスとタルボサウルスの幼体MPC-D 107/7はほとんど同じ大きさであるが、両者には形態学的に異なる点がいくつかある。頭骨の細かい形質が2点と、セレノらの報告によるとラプトレックスでは腸骨の寛骨臼の背方に垂直な稜がないことである。腸骨の垂直な稜はティラノサウルス上科の共有派生形質で、重要な点である。Tsuihiji et al. (2011) は、白亜紀前期の遼寧省という時代と産地をふまえた上で、これらの形態学的な差異から、暫定的にラプトレックスはタルボサウルス・バタールの幼体ではないだろうと述べている。
これについてホーナーらの論文では、白亜紀前期の中国遼寧省という点は既に根拠が否定されていること、ホーナー側の研究者がラプトレックスの標本を実際に観察したところ、腸骨にはかすかに垂直な稜がみとめられたことを指摘している。頭骨の2つの細かい形質については、Tsuihiji et al. (2011)も同種内の変異である可能性も否定できないとしている。これらのことからホーナーらは、ラプトレックスはタルボサウルス・バタールとは限らないが、タルボサウルス属か近縁の大型ティラノサウルス類の幼体である可能性が高いとしている。
産地が不明であることと幼体であることから、ラプトレックスを完模式標本として用いることには問題がある。特に、生息年代がわからない標本は他の標本と正確に比較することができない。また幼体と成体では形態が非常に異なることはよく知られているので、ラプトレックスが本当に新種だとしても、他の種類の成体と比較することはできない。他に有力な根拠が示されない限り、ラプトレックスは疑問名nomen dubiumと考えるべきである、とホーナーらはまとめている。
参考文献
Fowler DW, Woodward HN, Freedman EA, Larson PL, Horner JR (2011) Reanalysis of ‘‘Raptorex kriegsteini’’: A Juvenile Tyrannosaurid Dinosaur from Mongolia. PLoS ONE 6(6): e21376. doi:10.1371/journal.pone.0021376
Tsuihiji T, Watabe M, Tsogtbaatar K, Tsubamoto T, Barsbold R, et al. (2011) Cranial osteology of a juvenile specimen of Tarbosaurus bataar from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. J Vert Paleontol 31: 497-517.
Copyright 2011 Fowler et al.
コメント ( 3 ) | Trackback ( 0 )
特別展「新説・恐竜の成長」
大阪市立自然史博物館の特別展「新説・恐竜の成長」に行ってきました。
前半のメインはトリケラトプスの成長段階で、これはさすがに圧巻でした。眼の上の角の向き、フリルの長さ、縁後頭骨(フリルの縁にある三角形の骨)の状態、鼻の角と鼻骨の癒合などの比較展示です。DVD映像も使って子どもたちにもわかりやすく伝える工夫がされていました。
トリケラトプスの角やフリルは、「同種(内)の識別」のためという結論ですが、従来の「防衛のため」や「繁殖行動での雌雄間のディスプレイのため」という考えを「違います。」と完全否定してしまっていたのが気になりました。大丈夫かなあ。トリケラトプスが角を使えないでどうするのだろう。また雌雄ともフリルはあるが、模様が異なったりしないのだろうか。

ヒパクロサウルスとパキケファロサウルスの成長段階は中継ぎの位置にありました。この「ドラコレックス」からドーム状の成体への変化もかなり劇的ではあります。

後半の広いスペースの中央にはティラノサウルス動刻がいて、動いたり吠えたりしているので親子連れは釘付けになります。ロボットがガキんちょ、もといお子様を引きつけてくれるおかげで、化石・骨格の前はむしろ空いていたりするので、良しとしましょう。恐竜の動刻も年々進化しているらしく、かなり複雑な動きをしています。

それにしても、世界最大のティラノサウルス頭骨実物化石MOR008の前は妙に空いていました。あまり詳しい解説パネルがなかったせいか?


(スーの発見までは)最も保存の良いティラノサウルス骨格(だった)ワンケル。若い夫婦が、「やっぱりスーに抜かれたんだ」といいながら立ち去っていきました。しかし、初めて見る標本はやはり感動しますね。頭骨も保存がよく、立体感を演出するスポット照明が効いている。
ティラノサウルスについては、ホーナー博士の持論であるスカベンジャー説が展開されているのですが、スカベンジャーでもありハンターでもあったという視点が欠けていると感じました。なぜ二者択一なのでしょうか。大型の成熟個体がもっぱら屍肉食であったとしても、幼体や亜成体がハンターであることを認めるならば、ティラノサウルスは強力な捕食者であるという表現に問題ないと思うのですが。
全体としてこの企画は、「世界最大のなんたら」といったものよりは、はるかに学術的なテーマが明確であり、教育的プログラムもしっかりしている。ただそれだけに、特定の研究グループの学説のプロパガンダ色が感じられなくもない、という感じでしょうか。

今回のワンケル=MOR555Rは新しい復元キャストのようです。比較のため、日本在住の豊橋市のワンケル君を並べてみます。やはり新しいワンケルの方が洗練された感じがするかな。頭骨だけのは旧バージョンと同じ赤茶色でした。まあ頭部を下げている方が、顔がよく観察できて助かる。
前半のメインはトリケラトプスの成長段階で、これはさすがに圧巻でした。眼の上の角の向き、フリルの長さ、縁後頭骨(フリルの縁にある三角形の骨)の状態、鼻の角と鼻骨の癒合などの比較展示です。DVD映像も使って子どもたちにもわかりやすく伝える工夫がされていました。
トリケラトプスの角やフリルは、「同種(内)の識別」のためという結論ですが、従来の「防衛のため」や「繁殖行動での雌雄間のディスプレイのため」という考えを「違います。」と完全否定してしまっていたのが気になりました。大丈夫かなあ。トリケラトプスが角を使えないでどうするのだろう。また雌雄ともフリルはあるが、模様が異なったりしないのだろうか。
ヒパクロサウルスとパキケファロサウルスの成長段階は中継ぎの位置にありました。この「ドラコレックス」からドーム状の成体への変化もかなり劇的ではあります。
後半の広いスペースの中央にはティラノサウルス動刻がいて、動いたり吠えたりしているので親子連れは釘付けになります。ロボットがガキんちょ、もといお子様を引きつけてくれるおかげで、化石・骨格の前はむしろ空いていたりするので、良しとしましょう。恐竜の動刻も年々進化しているらしく、かなり複雑な動きをしています。
それにしても、世界最大のティラノサウルス頭骨実物化石MOR008の前は妙に空いていました。あまり詳しい解説パネルがなかったせいか?
(スーの発見までは)最も保存の良いティラノサウルス骨格(だった)ワンケル。若い夫婦が、「やっぱりスーに抜かれたんだ」といいながら立ち去っていきました。しかし、初めて見る標本はやはり感動しますね。頭骨も保存がよく、立体感を演出するスポット照明が効いている。
ティラノサウルスについては、ホーナー博士の持論であるスカベンジャー説が展開されているのですが、スカベンジャーでもありハンターでもあったという視点が欠けていると感じました。なぜ二者択一なのでしょうか。大型の成熟個体がもっぱら屍肉食であったとしても、幼体や亜成体がハンターであることを認めるならば、ティラノサウルスは強力な捕食者であるという表現に問題ないと思うのですが。
全体としてこの企画は、「世界最大のなんたら」といったものよりは、はるかに学術的なテーマが明確であり、教育的プログラムもしっかりしている。ただそれだけに、特定の研究グループの学説のプロパガンダ色が感じられなくもない、という感じでしょうか。
今回のワンケル=MOR555Rは新しい復元キャストのようです。比較のため、日本在住の豊橋市のワンケル君を並べてみます。やはり新しいワンケルの方が洗練された感じがするかな。頭骨だけのは旧バージョンと同じ赤茶色でした。まあ頭部を下げている方が、顔がよく観察できて助かる。
コメント ( 15 ) | Trackback ( 0 )
巨大ワニと恐竜の世界
大阪大学総合学術博物館の「巨大ワニと恐竜の世界」に行きました。
特別展示の会場は、限られたスペースに展示物を詰め込んだ感じでした。そのせいか、三畳紀のコーナーには恐竜エオラプトル、リンコサウルス類スカフォニクス、単弓類はいましたが、三畳紀のワニ類がいなかったような。このあたりを詳しくすると2010年の繰り返しになってしまうということか。写真パネルだけでもラウイスクス類などの姿が欲しかった気がしました。(次のジュラ紀のコーナーでは、ゴニオフォリスが既にワニらしい形態になっていると書かれているので、比較のためにも。)

受付の横の壁面にいきなりマチカネワニ。これはいいですね。

学問的に最も重要なのは、おそらくこれでしょう。現生種でマチカネワニと最も近縁なマレーガビアル。マレーガビアルとインドガビアルは全く別ものなのですね。マレーガビアルは吻が細長いが、二等辺三角形であることがよくわかりました。マレーガビアルはクロコダイル上科の中のトミストマ亜科の唯一の現生種。マチカネワニは最初、マレーガビアルの一種として記載されたが後に別属となり、さらに最近の研究で、トミストマ亜科に含まれマレーガビアルと最も近縁であることが確認されたということです。
マチカネワニの化石は下顎の先端が欠けていたが、これは病理的なもので治癒した痕跡があることから、この状態でしばらく生きていた。昔バナナワニ園かどこかで、下顎の先端が2つに裂けても平気で泳いでいるワニを見たことがありますが、ああいう感じだったのですね。

マチカネワニ生体復元模型(徳川さんの作品)。立体的なポーズが良い。
特別展示の会場は、限られたスペースに展示物を詰め込んだ感じでした。そのせいか、三畳紀のコーナーには恐竜エオラプトル、リンコサウルス類スカフォニクス、単弓類はいましたが、三畳紀のワニ類がいなかったような。このあたりを詳しくすると2010年の繰り返しになってしまうということか。写真パネルだけでもラウイスクス類などの姿が欲しかった気がしました。(次のジュラ紀のコーナーでは、ゴニオフォリスが既にワニらしい形態になっていると書かれているので、比較のためにも。)
受付の横の壁面にいきなりマチカネワニ。これはいいですね。
学問的に最も重要なのは、おそらくこれでしょう。現生種でマチカネワニと最も近縁なマレーガビアル。マレーガビアルとインドガビアルは全く別ものなのですね。マレーガビアルは吻が細長いが、二等辺三角形であることがよくわかりました。マレーガビアルはクロコダイル上科の中のトミストマ亜科の唯一の現生種。マチカネワニは最初、マレーガビアルの一種として記載されたが後に別属となり、さらに最近の研究で、トミストマ亜科に含まれマレーガビアルと最も近縁であることが確認されたということです。
マチカネワニの化石は下顎の先端が欠けていたが、これは病理的なもので治癒した痕跡があることから、この状態でしばらく生きていた。昔バナナワニ園かどこかで、下顎の先端が2つに裂けても平気で泳いでいるワニを見たことがありますが、ああいう感じだったのですね。
マチカネワニ生体復元模型(徳川さんの作品)。立体的なポーズが良い。
コメント ( 3 ) | Trackback ( 0 )