獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2019年2月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | |||||||
| 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| 10 | 11 | 12 | 13 | 14 | 15 | 16 | ||
| 17 | 18 | 19 | 20 | 21 | 22 | 23 | ||
| 24 | 25 | 26 | 27 | 28 | ||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
マジュンガサウルスの成長
Copyright 2016 Ratsimbaholison et al.
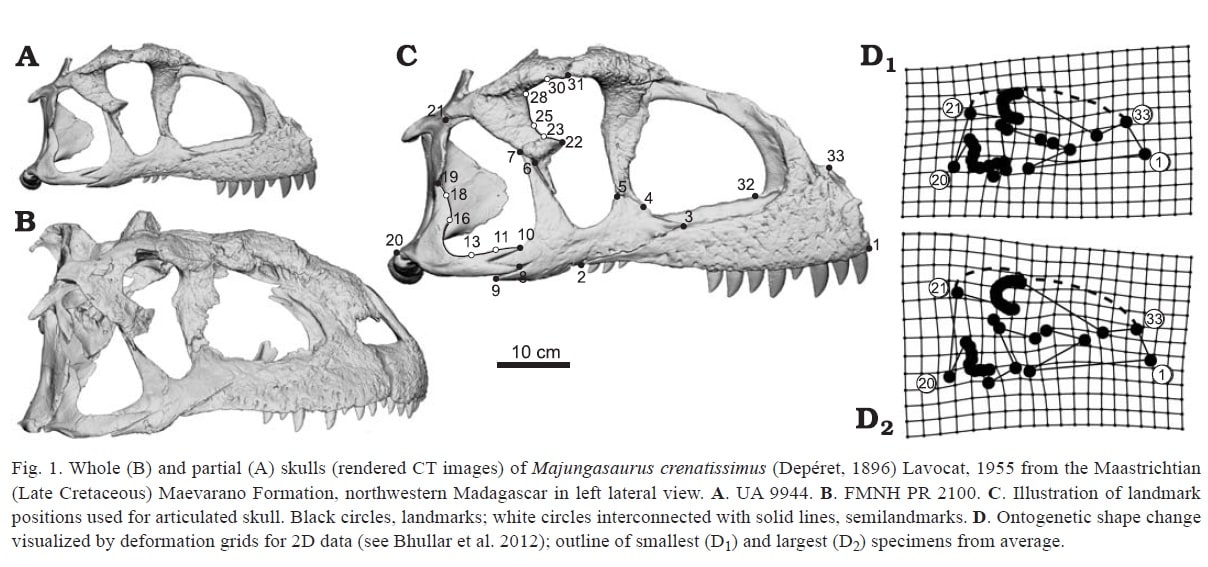
アベリサウルス類は獣脚類の中でも短く、丈の高い特徴的な頭骨をしている。しかしアベリサウルス類の化石はほとんどが一個体であり、種内変異や成長過程の研究はされていなかった。その中にあってマジュンガサウルスは近年、複数の個体の頭骨が発見されていることから、アベリサウルス類で初めて、成長過程の研究が可能となった。そこでRatsimbaholison et al. (2016) は、マジュンガサウルスの頭骨の成長過程を、幾何学的形態計測geometric morphometrics という手法で解析した。
幾何学的形態計測では、標本の形態をコンピューター上に取り込んでデジタル化し、異なる形態の間で対応づけられるような代表的な計測点(ランドマーク)を定める。そして座標変換のように形態を移行させ、変形した座標の形(deformation grid)が表示される。この方法はすでに、鳥類の頭骨が祖先の獣脚類の頭骨から幼形進化によって生じたことを示す研究や、多数の獣脚類の頭骨を定量的に比較した研究などに用いられている。
頭骨の要素を含むマジュンガサウルスの部分骨格は8個体分あるが、それぞれ保存された骨が異なるので、個々の骨について比較できるのは多くて5個体分くらいである。例えば前上顎骨は5個、上顎骨は4個、涙骨は2個、後眼窩骨は3個、頬骨は3個、方形骨は5個という具合である。アロサウルスなどと比べるとずっと少ないが、アベリサウルス類としては最も多い。
成長過程における前上顎骨の主な形態変化は、鼻骨突起と外鼻孔の下縁の角度が小さくなることと、鼻骨突起の長さの増大であった。
上顎骨の主な形態変化は、上顎骨体の前方部分の高さの増加と、腹側縁のS字状カーブの湾曲が減少することであった。また定性的には、上顎骨の外側面の粗面(彫刻)が成長とともに発達していた。
最も顕著な形態変化は、眼窩の周りや側頭部にみられた。涙骨では、涙骨全体の高さの増加、涙骨体の大きさの増加、眼窩の縁の周長の減少がみられた。眼窩の縁の減少は、骨の蓄積とともに眼窩下突起suborbital processの発達とも関係している。また涙骨の前方突起rostral ramusは、幼体では前方を向いているが、成体では前腹方にカーブしている。涙骨は2個しかないので、これが本当に成長に伴う変化なのか、あるいは種内変異なのか確認するためには、もっと多くのサンプルが必要であるという。
後眼窩骨では、成長とともに背側縁の長さの減少と、涙骨にみられたのと同様に眼窩の縁の減少が観察された。頬骨では、涙骨突起の高さの増加と、頬骨全体の長さの増加が観察された。また同時に、頬骨と方形頬骨の関節面の高さも増加していた。
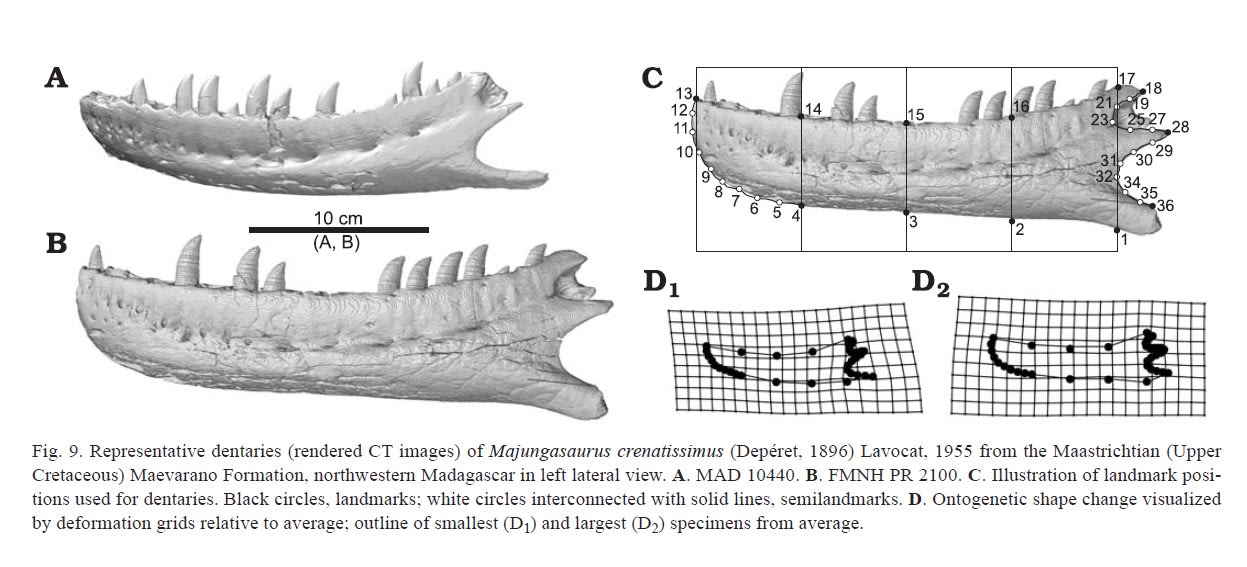
方形骨の主な形態変化は、後方の凹みconcavityの相対的減少と、方形骨軸の背側半分の高さの増大であった。歯骨の主な形態変化は、全体的な高さの増加と、歯骨の前腹側端が角ばった形から丸い形に変化することであった。上角骨については、とくに顕著な形態変化はみられなかった。
著者らは個々の骨の他に、3つの部分的に関節したマジュンガサウルスの頭骨(推定42 cm から53 cm)についても比較している。頭骨全体の変形座標をみると、成長に伴ういくつかの形態変化がわかった。まず、頬骨の涙骨突起の高さの増加と、頬骨の前方部分の前後長の増加が起きている。同時に頬骨と方形頬骨の関節面も増大している。また、大型の個体では頬骨の方向が回転しており、頬骨と上顎骨の関節面が、頬骨と方形頬骨の関節面よりも背側にくるように変化している。つまり幼体では水平だったのが、成体では後腹方―前背方の方向になっている。
2番目に、成長とともに眼窩の大きさは、頭骨のサイズに比べて相対的に小さくなっている。眼窩の下半分は幅が狭くなり、眼窩の上半分は直径が小さくなっている。
3番目に、頭骨の側頭部は高さが増大し、後眼窩骨の鱗状骨突起と方形頬骨の方形骨突起の間の高さが増加している。
結局、成長に伴って頭骨の丈が高くなるということであり、その点はよくあるパターンに思える。吻の部分では上顎骨の一部が高くなるだけであるが、眼窩の周りから側頭部にかけては全体に丈が高くなっているという。ただし今回、幼体の鼻骨がないために鼻骨の変化は観察できていない。もし鼻骨があれば、成長とともに盛り上がった形になるとか、吻の部分ももっと貢献しているという結論になるかもしれない。
獣脚類の頭骨は一般に、胚・新生児・幼体に比べると、成体では前後に長くなる傾向がある。コエロフィシスやコンプソグナトゥス類などではそうである。しかし、いわゆる大型獣脚類では、系統にかかわらず、途中から頭骨の丈が高くなるように変化するという。ティラノサウルス類の亜成体から成体への変化などがそうである。今回のマジュンガサウルスの研究から、アベリサウルス類の頭骨もこのような大型獣脚類のパターンを示すことがわかった。
感想として気になる点の一つは、上顎骨のラインである。上顎骨の腹側縁のS字状カーブは、成体の方が湾曲の程度が小さくなっているようにみえた。著者らはこれを、成長による変化とは関係ない個体変異の例としてあげているが、なぜそう言い切れるのだろうか。確か、ティラノサウルス類では幼体の方が直線に近く、成体の方が強くS字状にカーブする。それとは逆なので成長に伴う変化とは言いにくかったのか。しかし、アベリサウルス類ではそれが正常な成長パターンということはないだろうか。アベリサウルス類では成長の途中で早めにS字状カーブに達し、その後は湾曲が減少するように変化するのかもしれない。カルノタウルスのような極端な場合はもはやS字状ではなくなって、弧状になっている。
もう一つ気になるのは、歯骨のラインである。前腹側の“おとがい”のような部分は、一般的には成体の方が発達していそうに思えるが、マジュンガサウルスでは早めに“おとがい”が形成されて、成体では全体に膨らんで丸みを帯びるようになっている。これは、他の獣脚類でもみられることだろうか。
またチェナニサウルスの論文にあったように、歯骨の腹側縁はカルノタウルスなどではカーブしているが、マジュンガサウルスではまっすぐである。成体では確かにそうなっている。ところがなんと、マジュンガサウルスの幼体では腹側縁がカーブしているようである。これは意外だった。チェナニサウルスの論文には、ブラキロストラのようにカーブしている方が派生的にみえるが、アベリサウルス類ではその方が基盤的である可能性もある、とあった。それに関連している。
参考文献
Ratsimbaholison, N.O., Felice, R.N., and O’Connor, P.M. (2016). Ontogenetic changes in the craniomandibular skeleton of the abelisaurid dinosaur Majungasaurus crenatissimus from the Late Cretaceous of Madagascar. Acta Palaeontologica Polonica 61 (2): 281–292.
コメント ( 0 ) | Trackback ( 0 )
アベリサウルス類の脳はティタノサウルス類の夢をみるか
先日の古生物学会で聞いてきた小脳片葉で思い出した。脳エンドキャストの形状から機能についてどこまで論じられるのかという問題である。
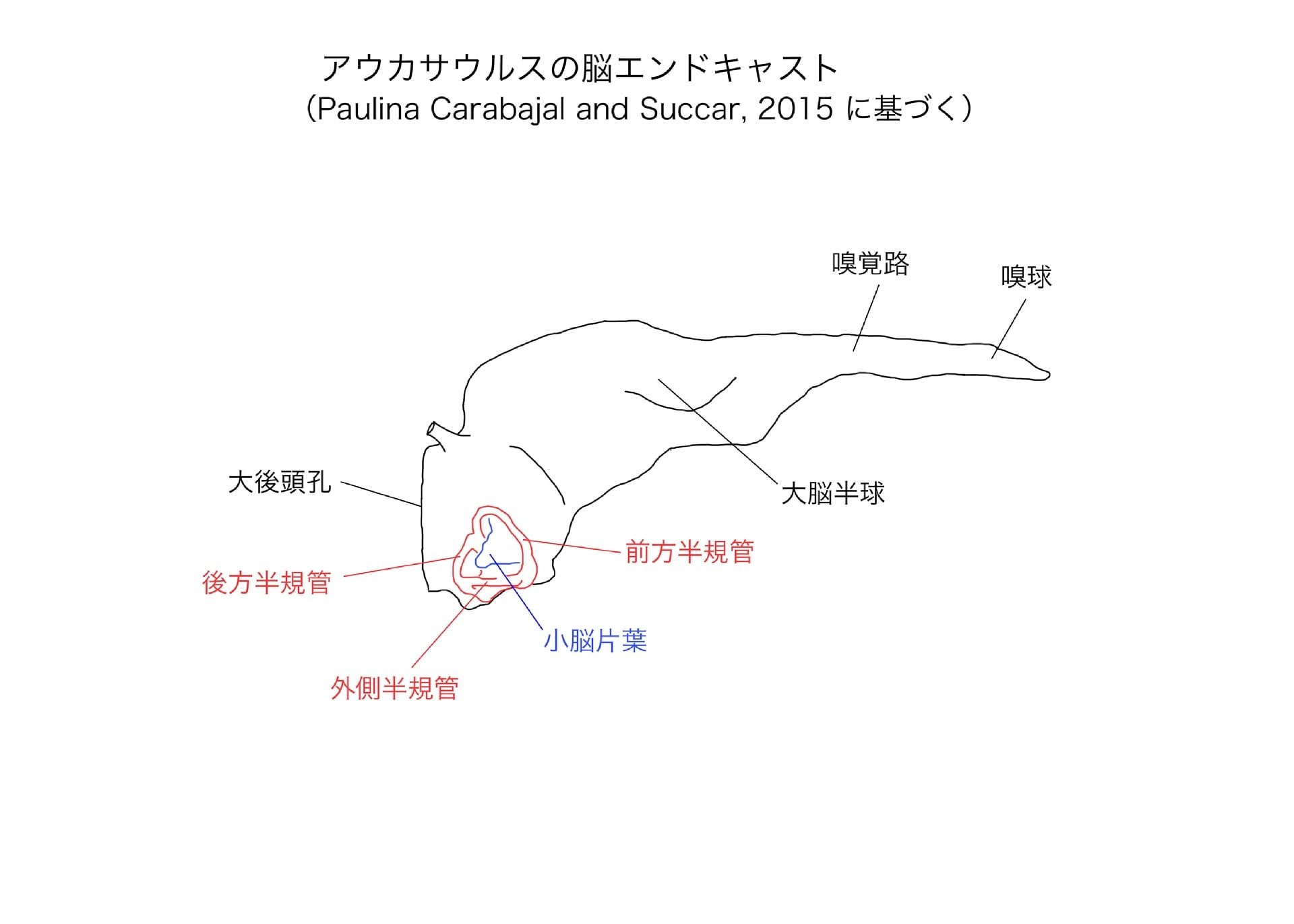
アベリサウルス類の脳函といえばPaulina Carabajal博士で、カルノタウルス、アベリサウルス、アウカサウルスの脳について研究している。アウカサウルスについては、脳函の腹側が大きく破損して内腔が露出していたが、CTスキャンによって脳エンドキャストを再現することができ、内耳の3次元構造も得られた。
アウカサウルスの脳をマジュンガサウルス、インドサウルス、ケラトサウルスと比較してみると、前脳、中脳、後脳のプロポーションは事実上これらの種類と同じで、基本的な形態はよく似ていた。
また内耳迷路(平衡覚器官)では、前方半規管が後方半規管より大きく、外側半規管が最も小さいが、これはほとんどの獣脚類と同様であった。外側半規管を水平にすると、アウカサウルスの頭蓋天井は水平に保たれている。これもマジュンガサウルスやケラトサウルスと同様で、基盤的な獣脚類に共通している。多くのコエルロサウルス類では頭蓋が下向きになる。
アウカサウルスの脳では、小脳片葉の部分がマジュンガサウルスよりも大きかった。小脳片葉は、頭、頸、体の動きに伴う眼球運動の調節(動眼反射)を司り、機敏な頭の動きに依存する動物種で大きく発達する傾向がある。そのためアウカサウルスはマジュンガサウルスよりも、すばやい頭の動きと動眼反射には優れていたかもしれないという。
しかし一方で、外側半規管の大きさは、マジュンガサウルスの方がアウカサウルスやケラトサウルスよりも大きく張り出していた。半規管が長い方が一般に感覚が鋭敏と考えられるので、頭を左右に振る運動については、マジュンガサウルスの方が感覚が鋭敏だったかもしれないという。
想像を逞しくすれば、アウカサウルスの方が高速で獲物を追跡することが多く、そのために動眼反射が発達したと思いたいところだが、それなら外側半規管が小さいのはどうなのか。一方マジュンガサウルスは、外側半規管で対応したということか。すばやい頭の動きに対して異なる適応の仕方をしたのかもしれない。
なんだか煮え切らないようだが、種間の変異として細かい違いはあっても、そこまで機能に反映していたとは言い切れないのではないだろうか。ある部分が同じ容積でも機能に差がある場合もあるだろうし、容積に差があっても機能は同じこともあるだろう。ニューロンの密度を調べたわけでも、感覚生理学的な実験をしたわけでもない。データに対してこのような考察を書くのは正しいが、機能については「可能性がある」くらいのものだろう。
参考文献
Paulina Carabajal, A. and Succar, C. (2015) The endocranial morphology and inner ear of the abelisaurid theropod Aucasaurus garridoi. Acta Palaeontologica Polonica 60 (1): 141–144.
コメント ( 0 ) | Trackback ( 0 )
チェナニサウルス(白亜紀末のモロッコのアベリサウルス類)
モロッコの大型アベリサウルス類といっても、過去の記事で取り上げた大腿骨頭と同じものではない。以前のはセノマニアンで、今回はマーストリヒティアンである。モロッコでは、白亜紀後期の初めセノマニアンのKem Kem bedからはアベリサウルス類の化石が知られていたが、白亜紀末期マーストリヒティアンからは分離した歯しか知られていなかった。アフリカ全体としても、初めてである。これまで白亜紀末(カンパニアンとマーストリヒティアン)のアベリサウルス類は、南アメリカ、マダガスカル、インド、パキスタン、ヨーロッパからは見つかっていたが、アフリカ大陸からは見つかっていなかった。今回、断片的ながら他のアベリサウルス類とは異なる特徴をもつ標本が発見されたので、命名記載されたものである。(最新のケラトサウリアの系統解析(Delcourt, 2018)ではアベリサウルス科から外れてケラトサウルス科に入っているが、ここではアベリサウルス科のカテゴリーに入れておく。)
チェナニサウルスは、白亜紀末マーストリヒティアン後期に北アフリカのモロッコに生息したアベリサウルス類で、2017年に記載された断片的な化石である。チェナニサウルス・バルバリクスChenanisaurus barbaricus の属名は産出地であるリン鉱山 Sidi Chennaneに、種小名はモロッコの古名バルバロス(バーバリー)に由来する。
チェナニサウルスのホロタイプ標本は、下顎の歯骨の前半部分である。これだけで、どうしてアベリサウルス類とわかるのだろうか。また、新種とされる特徴は何だろうか。
チェナニサウルスの特徴は、大型のアベリサウルス類(推定全長7~8 m)で下顎は比較的丈が高く、側面から見て歯骨がカーブしている、側面の溝とそれに伴う孔が歯骨の上方に位置する、下顎結合は太く、下顎結合の前縁は側面から見て垂直である、などである。
歯槽のサイズと比較して、歯骨は非常に丈が高いことから、比較的短く丈の高い下顎をしていたと考えられる。そのプロポーションはカルノタウルスやエクリクシナトサウルスと似ているが、丈の高さについてはもっと極端かもしれないという。保存された背側縁と腹側縁はカーブしており、ブラキロストラのアベリサウルス類と同様に、下顎の腹側がカーブしていたと考えられる。それに対してマジュンガサウルスでは歯骨の腹側縁はまっすぐである。
歯骨の側面の後方には、長軸方向の深い溝grooveがある。この溝の背方には一連の神経血管孔があり、それらの孔は背方の浅い溝sulcusに向かって開いている。同様の溝grooveと神経血管孔はカルノタウルスやマジュンガサウルスなど他のアベリサウルス類にもみられるが、これらの種類では溝が歯骨のもっと下方に位置している。それに対してチェナニサウルスでは、溝と神経血管孔が上方にあり、ゲニオデクテスと同じ原始的な状態であるという。(ゲニオデクテスは従来ケラトサウルス科とされてきたが、著者らは最も原始的なアベリサウルス類としている。)ケラトサウルスでは、溝は浅く上方にあるという。
また歯骨の側面には一連の窪みや吻合する稜があり、顕著な彫刻sculptureをなしている。同様の特徴はカルノタウルスやマジュンガサウルスなど派生的なアベリサウルス類にもみられる。ゲニオデクテスにはない。
内側面をみると、他のケラトサウリアと同様に歯間板が癒合している。歯間板は前方では非常に丈が高いが、後方へ行くにつれて急速に小さくなっている。丈の高い歯間板はアベリサウルス類の共有派生形質で、カルノタウルスやマジュンガサウルスのような派生的な種類にも、原始的なアベリサウルス類ゲニオデクテスにもみられるという。歯間板の表面には浅い溝(条線)がある。これはマジュンガサウルスなどにもみられる。
背側から見ると歯骨は外側に反っている。下顎結合の部分は左右の歯骨が鈍角をなすようになっている。背側から見て左右の歯骨は幅広いU字形をなすという。
10個の歯槽が保存されており、3個の歯冠と4個の歯根がある。歯冠は丈が高く、前縁は強く凸型にカーブしており、後縁は比較的まっすぐである。まっすぐな後縁はアベリサウルス類の派生形質で、カルノタウルスやマジュンガサウルスでは歯列全体にみられるが、ゲニオデクテスでは前上顎骨歯と前方の歯骨歯にのみみられるという。
Longrich et al. (2017) の系統解析では、南米の進化したアベリサウルス類(ブラキロストラ)のクレードと、インド・マダガスカル・ヨーロッパのアベリサウルス類(マジュンガサウリナエ)のクレードが再現されたが、チェナニサウルスはそれら2つのグループのどちらにも属さず、外側の基盤的な位置にきた。またアフリカのルゴプスも基盤的な位置にきている。ゲニオデクテスは最も基盤的なアベリサウルス類となっている。
チェナニサウルスは以下の多くの派生形質によりアベリサウルス科Abelisauridaeと認められる。
1)幅広いU字形の下顎結合
2)顕著な歯骨の側面の溝
3)大きな神経血管孔とこれらの孔から背方に伸びる溝sulcus
4)歯骨の側面後方にある顕著な彫刻sculpture
5)歯間板の表面に条線striationがある
6)歯間板は丈が高く、歯骨の内側面でかなり下方まで伸びている
7)亜直方形の歯槽
8)歯冠の前縁は強く凸型にカーブし、後縁はほとんどまっすぐである
9)歯冠の形が頂点から見て非対称で、歯冠の前外側面が強く凸型になっている
全体として、チェナニサウルスの歯骨の形態は、吻の長いマジュンガサウリナエよりも吻の短いブラキロストラのものに似ている。下顎はカーブしていて丈が高い。しかしカーブした下顎は、より基盤的な種類であるケラトサウルスやゲニオデクテスにもみられる原始形質であるといっている。つまり必ずしもブラキロストラに近縁とはいえないといっている。
側面の溝はマジュンガサウリナエでもブラキロストラでも腹方にあるが、チェナニサウルスでは背方にある。この点でチェナニサウルスはゲニオデクテスと似ている。今回の系統解析ではこのために両方のクレードの外側にきたと思われる。しかしチェナニサウルスや他のアベリサウルス類のデータが非常に限られていることを考えると、今回得られた系統関係は確かなものとはいえない。チェナニサウルスについても他のアベリサウルス類についても、もっと多くのデータが必要であるといっている。
確かに、普通に考えるとブラキロストラに入りそうにみえる。側面の溝の位置でゲニオデクテスの方に引っ張られたのだろうが、1つの形質でそうなるものなのだろうか。ちょっと気になるのは、今回の解析には地域の要素が入っていることで、アフリカ、インド・マダガスカル、南アメリカなどが形質データに含まれている。そのことも、今回のトポロジーに影響しているのではないだろうか。
ケラトサウリアの総説的な論文(Delcourt, 2018)ではケラトサウルス科に入っている。これもゲニオデクテスと共有する形質のためだろう。アフリカのアベリサウルス類については、もっと多くの標本が発見されないとまだまだというのが感想である。
参考文献
Longrich, N. R., Pereda-Suberbiola, X., Jalil, N.-E., Khaldoune, F. & Jourani, E. (2017) An abelisaurid from the latest Cretaceous (late Maastrichtian) of Morocco, North Africa. Cretac. Res. 76, 40–52.
Delcourt, R. (2018) Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific Reports 8:9730 DOI:10.1038/s41598-018-28154-x
コメント ( 0 ) | Trackback ( 0 )