獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年7月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | ||||
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| 13 | 14 | 15 | 16 | 17 | 18 | 19 | ||
| 20 | 21 | 22 | 23 | 24 | 25 | 26 | ||
| 27 | 28 | 29 | 30 | 31 | ||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
タウロヴェナトル

タウロヴェナトル・ヴィオランテイTaurovenator violanteiは、後期白亜紀セノマニアン中期からチューロニアン前期(Huincul formation)にアルゼンチンのリオ・ネグロ州に生息した大型のカルカロドントサウルス類で、2016年に後眼窩骨だけが記載されていたが、2024年に全身にわたる追加の標本を含めて改めて記載された。特に頸椎と前肢がよく保存されていて、カルカロドントサウルス類においてこれまで詳細が不明だった解剖学的特徴が得られている。Huincul formationではマプサウルス、メラクセスに次いで3番目のギガノトサウルス族となった。体のサイズはメラクセスとギガノトサウルスの中間である。
ホロタイプ標本は分離した右の後眼窩骨である。
参照標本は3-4 m2 の範囲から見つかった交連状態の骨格で、頭蓋の骨は頬骨、方形骨、口蓋骨、外翼状骨、上角骨、角骨、前関節骨、関節骨、角舌骨、3本の歯からなる。中軸骨格は環椎、頸椎3-6、後方の胴椎の神経弓、後方の胴椎の神経棘、胴椎の椎体、多数の頸椎と胴椎の肋骨、4つのガストラリアからなる。前肢の骨は上腕骨、橈骨、尺骨、中手骨I, II, III、指骨からなる。後肢の骨は大腿骨、遠位足根骨、中足骨I, II, III, V、趾骨からなる。
参照標本はホロタイプ標本から800 m離れた地点から発見された。両者には重複した骨はないが、産地が近く同じ層準で、どちらもギガノトサウルス族の特徴を示すことから同一種と考えられた。
他のカルカロドントサウルス類と区別されるタウロヴェナトルの特徴は、後眼窩骨の前方突起の後背側面に、角状で粗面のある突起をもつこと;後眼窩骨の腹側面に深いくぼみがあること;頬骨の方形頬骨と関節する2つの突起の間に、2つの孔があることなどである。その他、頬骨、方形骨、外翼状骨、大腿骨などの細かい形質からなっている。
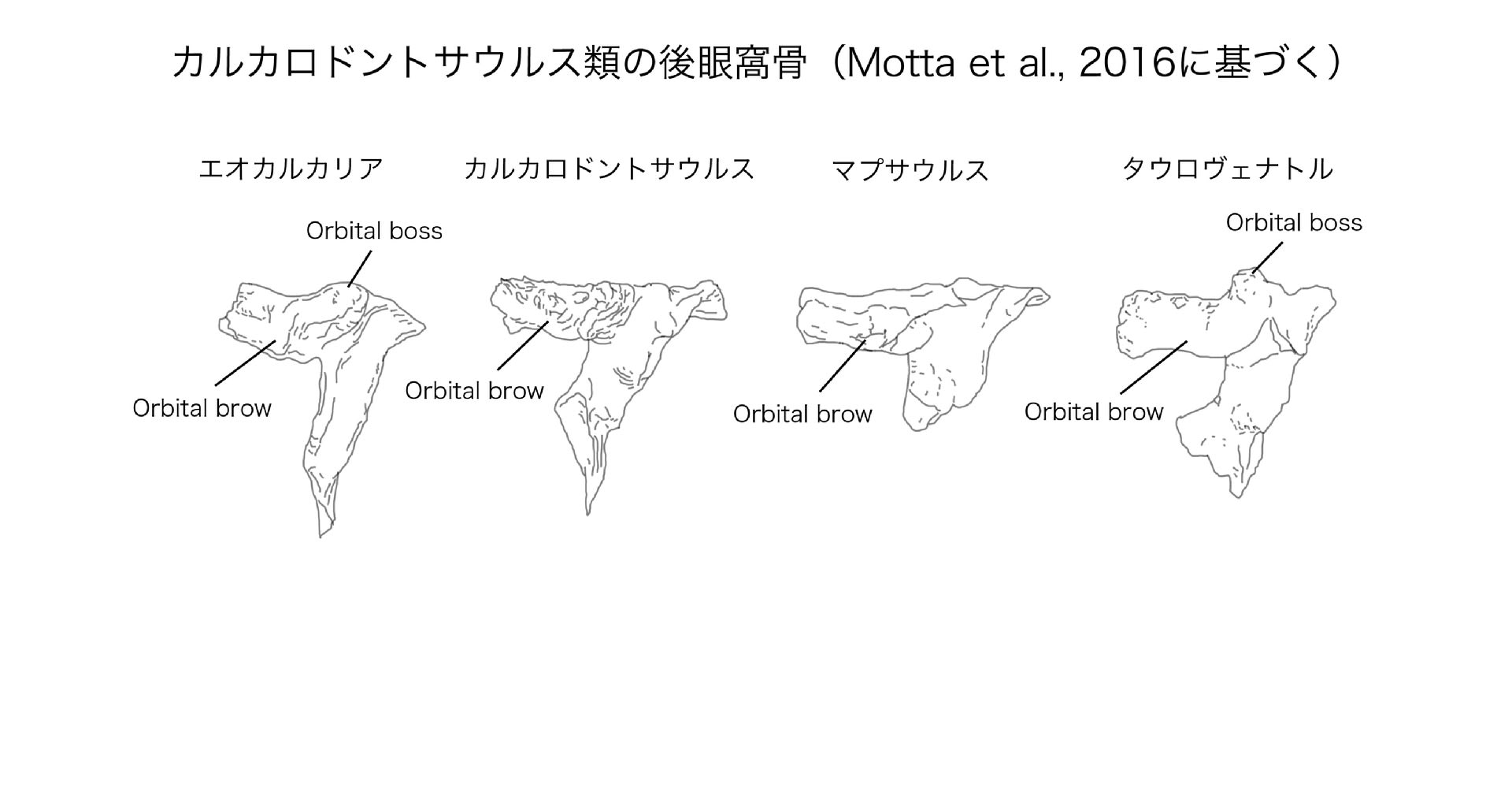
タウロヴェナトルの後眼窩骨には、他のカルカロドントサウルス類と同様に眼窩の上のひさし状の突起(orbital brow)がある。しかしタウロヴェナトルの特徴として、ひさし状の突起の後背方に、角状に突出した円錐形の突起(orbital boss)がある。基盤的なカルカロドントサウリアとされる(最近はあまりカルカロドントサウルス類に含まれない)エオカルカリアの後眼窩骨にはやや似た突起があるが、タウロヴェナトルほど突出していない。カルカロドントサウルスの後眼窩骨には細かい隆起はあるが、大きく突出するものはない。マプサウルスの後眼窩骨にも顕著な突起はない。
後眼窩骨の腹側突起が背腹に短く前後に幅広いこと、ひさし状の突起に対して垂直でなく前方に傾いていることは、派生的なカルカロドントサウルス類に共通した特徴である。

環椎以外の頸椎(C3-C6)をみると、他のカルカロドントサウルス類と同様に椎体は強く後凹型で、プレウロシール(側腔)は縦の骨壁で仕切られている。神経棘は高く、その頂上部は後方で幅が広がって背面から見て三角形であり、側面からみても上が広がった扇形となっている。その後方には深いくぼみがあり、次の頸椎の神経棘の前方部分がはまり込むようになっている。その結果C3-C6が関節した状態では、神経棘が重なり合った(imbricate)形となる。この状態はアクロカントサウルスのC3-C5 でも報告されており、また程度は小さいがこの傾向はアロサウルスやコンカヴェナトルにもみられるという。さらにタウロヴェナトルの頸椎ではエピポフィシスが上方を向いて大きく発達していることや、前関節突起と後関節突起の形状など、全体として頸椎を強化する特別な構造を形成している。著者らはこれを頸椎複合体cervical complexと呼んでいる。これはアクロカントサウルスにもみられ、また得られている情報からはギガノトサウルスやマプサウルスのような大型カルカロドントサウルス類にも共通している可能性がある。一方コンカヴェナトルやネオヴェナトルでは神経棘はより低く、エピポフィシスの形状も異なっている。
大型カルカロドントサウルス類は大きな頭骨をもち、強く後方に突出した球状の後頭顆をもつ。実はこれらの特徴はケラトプス類にもみられ、ケラトプス類では頸椎が一体化したsyncervicalという構造と関連しているとされている。球状の後頭顆は、頸椎に対して頭骨の可動範囲を大きくするのに役立つ。頸椎複合体は、頸椎を強化し筋肉の付着面を大きくすることができるが、個々の頸椎間の可動性は犠牲になっている。ケラトプス類と同じように、カルカロドントサウルス類の球状の後頭顆は、頸椎間の可動性が小さいことを補うように発達した可能性があるといっている。
もともと首が短いのに、なぜそこまでして頸椎を強化する必要があったのだろうか。これは、やはりかなり大型の植物食恐竜を獲物とすることと関係しているのではないか。想像をたくましくすれば、自分と同じくらいの大きさのティタノサウルス類の幼体などに咬みついた時に、獲物が死にもの狂いで暴れて、首が骨折して死亡する個体が続出したのかもしれない。そのような特殊な淘汰圧がかかったために、可動性を犠牲にしてでも頸椎を強化する方が有利となったのかもしれない。
論文中ではアロサウルス、アクロカントサウルス、メラクセス、タウロヴェナトルの前肢を並べているが、カルカロドントサウルス類の中でも派生的なギガノトサウルス族では最も前肢が縮小していることがわかる。タウロヴェナトルの前肢は、小さいことで知られるメラクセスの前肢よりもわずかに小さく、カルカロドントサウルス類の中で最小であるという。上腕が縮小する以上に前腕や手が縮小する点は、他の獣脚類と同様である。しかしメラクセスとタウロヴェナトルでは、手の骨のうち中手骨と末節骨が、末節骨以外の指骨よりも顕著に縮小している。つまり相対的に末節骨以外の指骨は長い。またこれらの指骨の関節面や筋付着部位はよく発達しており、十分に可動性のあるものであった。
ところで後眼窩骨以外はかなりメラクセスと似ているが、本当に新属新種でいいのだろうか。前肢の縮小パターンや後肢の第2趾の末節骨まで似ている。メラクセス属の別種ということはないだろうか。
参考文献
Rolando, A.M.A., Motta, M.J., Agnolín, F.L. et al. A new carcharodontosaurid specimen sheds light on the anatomy of South American giant predatory dinosaurs. Sci Nat 111, 56 (2024). https://doi.org/10.1007/s00114-024-01942-4
Motta MJ, Aranciaga-Rolando AM, Rozadilla S, Agnolín FE, Chimento NR, Brissón Egli F, Novas FE (2016) New theropod fauna from the Upper Cretaceous (Huincul Formation) of northwestern Patagonia, Argentina. New Mex Mus Nat Hist Sci Bull 71:231–253
コメント ( 0 ) | Trackback ( 0 )
タメリラプトル―名前を奪われた元祖カルカロドントサウルス―

ご存知の通りカルカロドントサウルスとは「サメの歯のトカゲ」という意味である。古生物ファンならよく目にする、モロッコ産のカルカロドントサウルスの歯は、確かにサメの歯に似ている。しかし、実はあれはサメの歯ほど左右対称ではなく、少し後方に傾いている。それに対して、元々のホロタイプのエジプト産カルカロドントサウルスの歯は、もっと左右対称でよりサメの歯と似ている。つまりモロッコ産カルカロドントサウルスをカルカロドントサウルス・サハリクスのネオタイプとしてしまったのはやや勇み足で、詳細に研究した結果、エジプト産とモロッコ産は別種と考えられた。よって本来はエジプト産がカルカロドントサウルス・サハリクスと呼ばれるべきだが、現在ではモロッコ産に基づいて、カルカロドントサウルス及びカルカロドントサウルス科という名称が学界ですっかり定着している。そのような諸般の事情に鑑みて、モロッコ産の名前を剝奪するようなことはせずそのまま認めて、逆にエジプト産の方を新属新種として命名した。そういう話である。
カルカロドントサウルス類の最初の化石記録は、アルジェリアのティミムン近郊で発見された2本の歯で、Deperet and Savornin (1925, 1927) によってメガロサウルス・サハリクスとして記載された。これを産出した地層は長年アルビアンとされてきたが、下部セノマニアンという説も出ている。
1931年にエルンスト・シュトローマーは白亜紀後期のエジプト産の化石に基づいて、最初のカルカロドントサウルス類の部分骨格を記載した。ホロタイプ標本SNSB-BSPG 1922 X 46は、エジプトのバハリア・オアシスで化石採集人マークグラフ氏が採集したもので、上顎骨、鼻骨、脳函、椎骨、部分的な恥骨と座骨、大腿骨、腓骨からなっていた。シュトローマーは上顎骨にあった歯の形態とアルジェリアの歯の類似性、つまりサメの歯に似ていることから、新属を提案しカルカロドントサウルス・サハリクスと命名した。これらの標本はすべて、第二次世界大戦のミュンヘン空爆によって失われた。現在まで残っている資料はシュトローマーの残した記載と図版、ベルリンに保管された脳函エンドキャスト、そしてこれまで記載されていない2枚の写真で、1つは左の上顎骨のもの、もう1つはミュンヘン古生物学博物館に展示されていた標本の写真である。
Sereno et al. (1996) はモロッコのケムケム層群からの比較的完全なカルカロドントサウルス類の頭骨を記載した。アルジェリアの歯およびエジプト産の標本との類似性により、カルカロドントサウルス・サハリクスとして記載し、2007年にはこれをネオタイプ標本とすることが提案された。しかし、エジプト産標本が失われていることもあり、エジプト産標本とモロッコ産標本の詳細な比較は、これまで行われてこなかった。only been done cursorilyという表現が面白くて、通りいっぺんの、ぞんざいなという意味である。ドイツの恐竜研究の金字塔であるシュトローマーの研究が、雑にしか検討されていないという憤りがあるのだろうか。
そこで今回、Kellermann et al. (2025) はシュトローマーの記載に加えて、再発見された未記載の写真資料と、現在のカルカロドントサウルス類の知見をふまえて再検討を行った。その結果、エジプト産標本はモロッコ産とは異なる形質を多数示し、新属新種タメリラプトル・マークグラフィTameryraptor markgrafiと命名された。
タメリラプトルの上顎骨には機能的な歯は保存されていないが、シュトローマーは左右の上顎骨に様々な成長段階の置換歯が保存されていることを観察している。1か所は歯を覆う骨の壁がプレパレーションで取り除かれて、歯冠の形が見えている。上顎骨にあるすべての置換歯は、高度に対称形で扁平であると記載されている。このように非常にサメの歯に似た歯冠は、他のどの獣脚類にもみられない。ただし興味深いことに、モロッコのケムケム産の分離した歯の断片と、マプサウルスとされる分離した歯(MCF-PVPH-108.103)にはこのような形がみられるという。モロッコやアルゼンチンにもこのような歯をもつカルカロドントサウルス類がいたのだろうか。
完全に萌出した歯がどのような形状だったかはわからない。さらに興味深いことに、エオカルカリアとされる上顎骨では、5番目の歯槽に保存された小さい置換歯は非常に対称形であるが、より成長した7番目の置換歯は少し後方に傾いている。Hanai and Tsuihijiによるタルボサウルスの歯の成長の研究では非常に小さいうちから後方に曲がっているので、歯の成長過程でより後方に曲がることが、カルカロドントサウルス類に特有の成長パターンであった可能性がある。しかし現在のところカルカロドントサウルス類の歯の成長の研究は行われていないため、これについては評価できない。
タメリラプトルと異なり、カルカロドントサウルス・サハリクスの上顎骨の歯はすべて、典型的な後方への屈曲を示し、歯の先端が基部の後半に位置している。観察されるタメリラプトルの置換歯はかなり大きいので、歯が完全に成長したとしても、このようなレベルの屈曲には達しないと思われる。
左右の鼻骨は全長にわたって大体同じ幅であるが、中央部が少しくびれている。この部分では内側の関節面が斜めになっているので、左右の鼻骨が正中線上で屋根状roof-like fashionに結合しており、正中で低い稜となっている。これはタメリラプトルに特徴的で、カルカロドントサウルスや他のカルカロドントサウルス類では左右の鼻骨の縫合線は平らであり、後方ではやや窪んでいたりする。
鼻骨の背面は大きな隆起のある粗面をなす。この粗面は前方では背内側部に局在すると記載されている。この点はカルカロドントサウルス、ギガノトサウルス、マプサウルス、メラクセスなどの他のカルカロドントサウルス類と同様である。特筆すべき鼻骨の特徴は約3㎝の角状の突起で、左右の鼻骨の正中側にある。ケラトサウルスの角ほど大きなものではないが、このような角は他のどのカルカロドントサウルス類でも知られていない。これは見たところ、カルカロドントサウルス類やティラノサウルス類の鼻骨にあるごつごつした隆起が特に発達したもののようにみえる。確かに正中にある稜状ではあるが、ケラトサウルスのトサカのように大きなものではなく、むしろアリオラムスの角のように小さく複数あってもよさそうな構造ではないだろうか。シュトローマーは角の後方にも切り株のような形の隆起をスケッチしている。
他のアロサウロイドと同様に、鼻骨の外側面に前眼窩窩が広がっている。タメリラプトルでは、この前眼窩窩が側面から見えている。このことは他の派生的なカルカロドントサウルス類と異なる。カルカロドントサウルス、ギガノトサウルス、マプサウルス、メラクセスでは、背側の棚状部分がオーバーハングして前眼窩窩は隠れている。
他のカルカロドントサウルス類と同様に、上顎骨の外側面にはごつごつした粗面が発達し、顕著な縦の稜と溝で覆われている。その発達の程度はギガノトサウルス、カルカロドントサウルス、メラクセスよりも著しい。前眼窩窩との境界には、強く発達した稜がある。シュトローマーによるとこの前眼窩窩の縁は、上顎骨の表面から3 cmも突出している。この稜の発達の程度は、カルカロドントサウルス・サハリクスと最も似ている。マプサウルス、メラクセス、ギガノトサウルスではもう少し発達が弱く、コンカヴェナトル、アクロカントサウルス、カルカロドントサウルス・イグイデンシスではさらに弱い。
タメリラプトルの脳エンドキャストは、他のカルカロドントサウルス類と大体似ている。カルカロドントサウルス・サハリクスとタメリラプトルを比較すると、タメリラプトルの方が相対的に大脳が大きい。嗅球と大脳の比率をみると、カルカロドントサウルスとギガノトサウルスは似ており、タメリラプトルの形はアクロカントサウルスと似ている。(それよりカルカロドントサウルス・イグイデンシスの嗅球の形が他のカルカロドントサウルス類とずいぶん違うようにみえるが、これは何なのだろう。)
カルカロドントサウルスの大腿骨というのは、このエジプト標本のことだったはずである。タメリラプトルの大腿骨は、骨幹の部分がつぶれていて直径の推定はできないとある。しかし近位端と遠位端は形がよく保存されている。大腿骨頭は、マプサウルス、メラクセス、ギガノトサウルス、ティランノティタンのような他のカルカロドントサウルス類と同様に、背内側を向いている。その程度はコンカヴェナトルやアクロカントサウルスよりも大きい。またタメリラプトルの大腿骨頭は、丸く背側に膨らんでいる点が他のカルカロドントサウルス類と異なっている。
小転子の下にトゲ状の付属転子accessory trochanterがある。このように顕著な三角形の付属転子はネオヴェナトルとアクロカントサウルスにのみみられ、アロサウルスやシンラプトルにも、他の派生的なカルカロドントサウルス類にもみられない。
系統解析の結果、タメリラプトルは派生的なカルカロドントサウルス科で、カルカロドントサウルス亜科(カルカロドントサウルスとギガノトサウルス族)よりは外側にきた。つまりカルカロドントサウルスよりは基盤的で、アクロカントサウルスよりは派生的な位置である。タメリラプトルがカルカロドントサウリア(コンカヴェナトル以上)と共有する形質は、上顎骨の歯間板が癒合している、前頭骨と頭頂骨が癒合している、大腿骨頭が背内側を向いている、などである。またカルカロドントサウルス科と共有する形質は、腸骨と座骨の関節がpeg and socket 構造をなすことである。
この研究の系統解析では、メトリアカントサウルス類とカルカロドントサウルス類が姉妹群になることが多いというのが面白い。アスファルトヴェナトルはアロサウルス科になったり、基盤的なアロサウロイドになったりしている。ちなみにメガラプトル類はほぼティラノサウロイドに含まれている。
参考文献
Kellermann M, Cuesta E, Rauhut OWM (2025) Re-evaluation of the Bahariya Formation carcharodontosaurid (Dinosauria: Theropoda) and its implications for allosauroid phylogeny. PLoS ONE 20(1): e0311096. https://doi.org/10.1371/journal.pone.0311096
コメント ( 0 ) | Trackback ( 0 )
ティランノティタン単品

ショップの全商品を見たわけではないが、ティランノティタンの全身復元のイメージはTシャツデザインしかなかったように見えた。もちろん公式ガチャにも入れてもらえなくてかわいそう。
ティランノティタンの完全なイメージはないので、描いてるうちにメラクセスにも似てくるのは仕方がない。しかしティランノティタンは南米大陸にカルカロドントサウルス類の覇権を確立した英雄であり、セノマニアンのギガノトサウルス、マプサウルス、メラクセスも偉大な祖先の墓参りに訪れたはずである。
コメント ( 0 ) | Trackback ( 0 )


中型ティタノサウルス類とギガノトサウルス族

合同展示会に来訪された方々、お買い上げ頂いた皆さま、ありがとうございました。
中型ティタノサウルス類は種類は特定せず、ディアマンティナサウルスのように14mくらいのものを想定。ギガノトサウルス族とは南米の大型カルカロドントサウルス類のグループで、ギガノトサウルス、マプサウルス、ティランノティタン、メラクセスが含まれるが、イメージしたのはギガノトです。
コメント ( 0 ) | Trackback ( 0 )

ハオロング風

最近は、PNSOやHaolonggoodの模型からヒントを得て絵を描いたりしているのがバレバレである。
熱帯の爬虫類といっても熱帯雨林のカメレオンなどと異なり、乾燥した平地の大型爬虫類としてはそれほど派手な色彩にはしないのが普通だろう。マンドリルのように顔だけ派手な色というのはありうるかもしれない。Haolonggoodは尾の先端がオレンジ色などになっているのがトレードマークであるが、手や足の裏が赤いのはやりすぎと感じた。
コメント ( 0 ) | Trackback ( 0 )
アクロカントサウルスは米国東部にも生息していた
アクロカントサウルスは前期白亜紀の終わりに米国に生息したカルカロドントサウルス類であるが、これまではテキサス、オクラホマ、ワイオミングの各州から見つかっていた。今回、東部大西洋岸のメリーランド州から発見された部分化石が、初めてアクロカントサウルスと同定された。これによりアクロカントサウルスは当時の北アメリカに広く分布していた可能性が高まった。
メリーランド州にはArundel Clay (Arundel formation)という前期白亜紀の地層があり、多くは断片的ながら恐竜化石が発見されてきた。1992年にUSNM 466054という獣脚類の部分化石が発見された。これは断片的だが交連状態の部分骨格で、第3胴椎の神経弓の一部、別の前方胴椎の神経弓の一部、2個の尾椎、大腿骨の近位部、脛骨の近位部と中央部、踵骨、いくつかの趾骨とその他の骨の断片からなる。前期白亜紀の米国東部では最も完全な獣脚類化石であるという。
USNM 466054は過去の研究で何度か取り上げられているが、いくつかの研究ではオルミトミムス類と推測されていた。それが今回はアロサウルス上科で、アクロカントサウルスの特徴を示すとなった。ずいぶん体形が異なるが、オルミトミムス類と考えられていたものが、なぜアクロカントサウルスになったのだろうか。
Weishampel and Young (1996) は「獣脚類の中で、足の爪が扁平で、脛骨に前方に突出した大きな稜をもつのはオルニトミムス類だけである」と述べている。しかし今回のCarrano (2023) によると、USNM 466054の脛骨突起(脛骨稜)cnemial crestは獣脚類としては特に大きいわけではないという。それは多くのオルニトミムス類と似ているが、多くのアロサウロイドやティラノサウロイドとも同じくらい似ている。つまりその形態は、基盤的なコエルロサウルス類や派生的なテタヌラ類(コエルロサウルス類以外の)に共通したものである。オルニトミムス類と他のテタヌラ類や基盤的コエルロサウルス類を区別できるような脛骨の特徴はほとんどないという。
同様に、USNM 466054の足の末節骨の形態は、基盤的なコエルロサウルス類や派生的なテタヌラ類に典型的なものである。多くの獣脚類で、足の第II-IV 指の末節骨は平らな腹側面をもち、断面が三角形である。一方オルニトミムス類の特徴は、末節骨の腹側面の内側縁と外側縁が外側に膨らんで、底が平らなテーブル状の形態となることである。このような特徴はUSNM 466054にはない。
さらに、USNM 466054は1つのアクロカントサウルスの固有派生形質をもつ。それは背側から見たとき、大腿骨頭が内側に向かって先細りになっていることである。この標本では肝心の大腿骨頭の先端が欠けているが、保存された部分だけで大腿骨頭の前縁と後縁が収束しているのがわかるという。これは他のアロサウロイドにはみられないが、アクロカントサウルスの他の標本と一致する。図をみると確かに他のアクロカントサウルスの標本と似ているようだ。ただし著者は、アクロカントサウルスの系統にはまだ知られていない多くの種がいたはずなので、この標本をアクロカントサウルス・アトケンシスとはせず、Acrocanthosaurus cf. A. atokensisとしている。
胴椎の神経弓と聞いて、神経棘か?と期待したが、全然違った。第3胴椎の神経弓といっても神経棘は欠けていて、基部の左半分のみである。椎体はきれいに外れているので、亜成体であることがわかる。
これらの神経弓が前後に短いことは、アロサウルス、アクロカントサウルス、マプサウルス、ティラノサウルスのような大型獣脚類の形態と一致する。前後に長いオルニトミムス類の胴椎とは異なる。さらに神経弓が前方に傾いていることは、アロサウルスとアクロカントサウルスの前方の胴椎にみられるが、他の大型獣脚類ではより垂直であるという。
参考文献
Carrano, M. T. First definitive record of Acrocanthosaurus (Theropoda: Carcharodontosauridae) in the Lower Cretaceous of eastern North America. Cretaceous Research, https://doi.org/10.1016/j.cretres.2023.105814
コメント ( 2 ) | Trackback ( 0 )
カルカロドントサウルスが獲物を持ち上げる力

Copyright 2015 Henderson & Nicholls “Double Death” という作品
過去に話題になったかもしれないが、私は最近知った。変わった研究があるものである。著者の一人はアーティストで、2頭のカルカロドントサウルスが竜脚類の亜成体を持ち上げている絵を作成した。それを見て実際にこんなことが可能なのか、という疑問から研究が開始された。筆頭著者はHendersonで、いろいろな動物の3次元デジタルモデルを作る人である。スピノサウルスの浮力や安定性についてもシミュレーションしている(「スピノサウルスは沈まない」の記事)。実際にカルカロドントサウルスはどのくらいの重さの獲物を持ち上げて、よろめかずに立っていられるのか、ということを真面目に研究した。
そのためにまず、コンピューター上でカルカロドントサウルスの3次元デジタルモデルを作成した。胴体についてはアロサウルスやアクロカントサウルスを参考にしている。例によって尾部は重く、前半身は含気性の骨や気嚢系があって軽いので、それぞれに適切な密度を与えている。その結果、カルカロドントサウルスのモデルは全長12.5 m、体重6.23 tとなった。一方の竜脚類の方は、カルカロドントサウルスと共存したレッバキサウルスが不完全なので、アルゼンチンのレッバキサウルス類リマイサウルスの3次元デジタルモデルを作成した。絵の中でカルカロドントサウルスが12.5 mとすると、竜脚類は10.9 m、1.91 t と推定された。
著者らは3つの観点から考察している。1)重心の移動、2)頭を持ち上げる力、3)顎でくわえる力、である。1)重心の移動については、カルカロドントサウルスのモデルの横断切片の重量を計算し、加算することで、重心の位置を決める。後肢を少し前後に開いた自然な立ちポーズで、重心は骨盤のすぐ前方にきた。獲物を口にくわえた場合、重心は前方に移動する。あまりにも重いものをくわえると、重心が前方に行き過ぎ、前のめりになって立っていられないわけである。その限界は重心が前に出した足の上にあるかどうかとした。
この計算式を解くと、1頭のカルカロドントサウルスが持ち上げて立っていられる荷重の上限は、2,510 kgとなった。2頭がかりだと5,020 kgとなり、最初の絵の竜脚類が1,910 kgなので、十分可能に思われた。つまり重心の移動という観点だけからは、最初の状況が可能にみえる。しかし実際には、首や顎の筋力の限界の方が小さいので、それらに制約されることがわかった。
2)頭を持ち上げる力については、獲物をくわえた頭全体を、首の背側にある軸上筋で支えるわけである。最も重要なのは後頭部に付くm.transversospinalis とm.complexusという2つの筋肉で、これらは大体頭頂骨の後部に付着する。片側のm.transversospinalis とm.complexusの断面積はアロサウルスで36.3 cm2 と 33 cm2と計算されている。カルカロドントサウルスの頭骨長はアロサウルスの2.26 倍であり、断面積は長さの増加の2乗に比例するので、2.26の2乗で5.125倍と推定された。また恐竜の筋力の推定値には幅があるが、ここでは40 N/cm2とされた。頭骨と頸椎が関節する後頭顆の位置を支点として、これらの首の筋肉は頭を持ち上げる(上に回転させる)ように働く。一方、頭自体の重量と口にくわえた獲物の重量は、頭を下げる(下に回転させる)ように働く。そこで
頭を持ち上げる力(モーメント)=頭自体の重量で下がる力+獲物の重量(最大荷重)で下がる力
という方程式を立てた。これを解くと、持ち上げられる獲物の重量の最大値は424 kgとなった。2頭がかりだと848 kgとなる。848 kg のリマイサウルスは8.29 mとなるので、最初の絵の3/4の大きさならなんとか可能という結果になった。
3)顎でくわえる力については、下顎と獲物の重量を、顎を閉じる筋肉(下顎内転筋)の力で支えることになる。アロサウルスの咬む力は歯列の前方で4,179 N、後方で6,809 Nと推定されているので、歯列の中央では5,494 Nとなる。下顎内転筋の断面積は長さの増加の2乗に比例する。頭骨長がアロサウルスの0.72 m からカルカロドントサウルスの1.63 mにスケールアップすると、下顎内転筋の断面積は2.26の2乗倍に増える。そして咬む力が断面積に比例するとすると、カルカロドントサウルスの咬む力は5,494 N x 2.26の2乗 = 2.82 x 10の4乗 N となる。これが最大の荷重(獲物の重量)と拮抗するという方程式を立てた。これを解くと獲物の重量の最大値は512 kgとなった。


Copyright 2015 Henderson & Nicholls
以上の結果から著者らは、より実際にありそうな絵を描きなおしている。最初の絵と同じくらいリアルだったらなお良かったが、このくらいの幼体なら持ち上げられそうな感じになった。ただしこの400-500 kgというのも上限なので、通常はずっと小さいものを持ち上げただろうと述べている。400 kgとするとウシは無理だがブタくらいなら持ち上げられるのかもしれないが、重量挙げのように一瞬の間だろう。
ずっと小さい獲物をくわえただろうという理由の一つとして、小さい物体は首、頭、顎の動きによってより簡単に扱うことができるということがある。多くの肉食爬虫類はinertial feeding という摂食方法をとる。これは小型の獲物を口にくわえて、すばやく首を振り、獲物を回転させて飲み込みやすい向きにするというものである。ティラノサウルスではinertial feedingの能力について研究されており、50 kgの肉塊をinertial feedingでくわえなおすことが可能らしい。カルカロドントサウルスでも同じような大きさの塊をinertial feedingで扱った可能性が高い。
感想として、元々の設定に無理があるのではないか。最初の絵は直感的に見て、これはちょっと重すぎて無理ではないかと思えた。そのことを、3次元モデルを作成して実証した研究ということになる。この2頭のカルカロドントサウルスが竜脚類を持ち上げている様子は、「公開処刑」を見せつけているように見えて、人間の発想に思える。(だから作品としてインパクトがあるのだろうが。)そもそも2頭の獣脚類が、それなりに重い竜脚類の体全体を地面から持ち上げる理由がない。ごく小さい幼体なら飲み込もうとして持ち上げる、または2頭で引っ張り合いをした結果持ち上がることはあるだろう。しかしそれなりに大きい獲物の場合は、獣脚類が持ち上げるのは飲み込める大きさに引きちぎった肉塊のはずである。獲物の体を地上に置いたまま解体し、ちぎっては食べ、ちぎっては食べる方が自然である。
参考文献
Donald M. Henderson, Robert Nicholls (2015) Balance and Strength—Estimating the Maximum Prey-Lifting Potential of the Large Predatory Dinosaur Carcharodontosaurus saharicus. THE ANATOMICAL RECORD 298: 1367–1375. https://doi.org/10.1002/ar.23164
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ |