獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年8月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | |||||||
| 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| 10 | 11 | 12 | 13 | 14 | 15 | 16 | ||
| 17 | 18 | 19 | 20 | 21 | 22 | 23 | ||
| 24 | 25 | 26 | 27 | 28 | 29 | 30 | ||
| 31 | ||||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ポーランドにおける三畳紀からジュラ紀にかけての恐竜時代の到来を表す消化管内容物化石
これは、同じ著者らによる2019年のスモックの糞化石の研究を、はるかに拡張して大きい仕事にまとめた感じである。三畳紀のポーランドに生息した全長5~6mの肉食主竜類スモックは、ティラノサウルスのように獲物の骨をかみ砕く頂点捕食者であり、ディキノドン類の骨とともに自分の折れた歯の断片も一緒に飲み込むという、獰猛で豪快な捕食者であった。大型のディキノドン類リソウィツィアや両生類などを含むこの時代の生態系の食物連鎖については、すでに再構成されていた。
今回の大規模な研究では、いわゆる糞石coprolitesのほかに嘔吐物が化石化した嘔吐石 regurgitalites、消化管が化石化した腸石cololitesも含めた消化物化石bromalitesをすべて集めている。ポーランド各地から532の消化物化石を集め、シンクロトロンマイクロCT による最新の高解像度画像解析、ガスクロマトグラフィーマススペクトルGC-MSなどによる化学分析などを駆使して、徹底的に解析している。
魚類の鱗や鰭を含む大型でらせん状の糞化石は大型の肺魚のもの、かみ砕かれた骨や歯を含む糞化石は肉食主竜類の頂点捕食者スモックのもの、ワニ形類の骨を含む大型獣脚類の糞化石、完全な甲虫などの昆虫を含む糞化石はシレサウルス、両生類や魚類を含む糞化石はラウイスクス類ポロノスクス、竜脚形類と思われる植物を含む糞化石、大型で魚類を含む糞化石はフィトサウルス類パレオリヌス、植物を含むディキノドン類の糞化石などが同定されている。
これらの化石をもとに食物連鎖を再構築し、三畳紀カルニアンからジュラ紀初期ヘッタンギアンにかけて5つの時代の生物相を解析した。その結果、始めは偽鰐類や獣弓類など恐竜以外の動物が優勢で、恐竜はほんのわずかな小型の種類だけであったのが、徐々に恐竜以外が衰退し、植物食の偽鰐類や獣弓類は竜脚形類や初期の鳥盤類で置き換えられ、その後獣脚類が大型化し偽鰐類に代わって頂点捕食者の地位を占めるようになった様子が裏付けられたという。
参考文献
Qvarnström, M., Vikberg Wernström, J., Wawrzyniak, Z. et al. Digestive contents and food webs record the advent of dinosaur supremacy. Nature (2024). https://doi.org/10.1038/s41586-024-08265-4
コメント ( 0 ) | Trackback ( 0 )
サウロスクスの機能形態学

Copyright 2023 Fawcett et al.
サウロスクスといえば、後期三畳紀アルゼンチンのイスキグアラスト層の動物相において、最強の肉食動物であり、ヘレラサウルスも逃げ出すほどの頂点捕食者のはずである。初期の小型の恐竜にとっては到底かなわない相手であり、後の時代の大型獣脚類に匹敵するパワーをもち、獣脚類のように丈の高い頭骨にはナイフのような鋭い歯がずらりと並び、咬む力も・・・と書きたいところである。
ところが最新の研究によると、サウロスクスの頭骨は全体としてはそれなりに頑丈であるが、一部弱い部分があり、咬む力は意外に弱くアロサウルスの半分程度であるという。これはかなりがっかりである。何かの間違いであってほしい。
Fawcett et al. (2023) はラウイスクス類で初めて、保存の良いサウロスクスの頭骨をretrodeformationという手法で完全に復元した。そして顎の筋肉の起始・付着面を同定し、顎を閉じる筋肉の断面積と力を算出した。有限要素解析で物を咬んだときの応力分布を視覚化し、すでにモデル化されているアロサウルスとサウロスクスの頭骨を比較した。さらに強い力を加えてそれぞれの頭骨を変形させるシミュレーションも行っている。
サウロスクスとアロサウルスの頭骨を比較すると、物体を咬んだ時の応力分布は頭骨の後半部では同じようなパターンを示した。しかし頭骨の前半部、特に口蓋の前方部分はアロサウルスの方がサウロスクスよりも強いという。サウロスクスの鋤骨は薄く、口蓋部分からの力を強く受けやすい。サウロスクスの頭骨では鋤骨、翼状骨、方形骨などが力学的に弱い部分であるという。
サウロスクスの咬む力は、歯列の前方で1015 N、後方で1885 Nと推定された。アロサウルスでは歯列の後方で3500 Nと推定されているので(Rayfield et al., 2001)、これはアロサウルスの半分程度ということになる。アロサウルス自体が全長7.5 mの動物としては弱いといわれていたので、サウロスクスの咬む力はかなり弱いという。もちろんティラノサウルスよりははるかに小さい。(この論文では片側の筋肉の力を表記している。)
現生ワニの中でサウロスクスと最も近いのは、インドガビアルの前方924 N、後方1895 Nである。インドガビアルも現生ワニ類の中では弱い種類であり、大体全長が2 mより大きいワニでは、これよりも強いという。現生ワニ類は咬む力が最も強い動物群であるが、系統が近いからといって同じように強いとは限らないという。
魚食のインドガビアルと同じではいかにも弱いように思える。一つの希望は、ラウイスクス類の中でサウロスクスは咬む力が弱い種類かもしれないということである。部分的なサウロスクスの下顎の前端は、背腹に広がっていないという。バトラコトムスやポストスクスの下顎の前端はもっと広がっている。つまり他のラウイスクス類はサウロスクスよりも咬む力が強いという可能性に期待したい。ラウイスクス類ではないかもしれないがポーランドのスモックは、骨片を含む糞化石から骨ごと砕いて食べたと考えられている。それとの関連はどうなのだろうか。
頭骨を変形させるシミュレーションについて、興味深いことをいっている。サウロスクスの頭骨は、側面から見て大体長方形に近い形である(上顎骨の下縁は下に凸)。この形だと前方で物を咬んだときに、吻を上に曲げるような変形を受けやすい。アロサウルスの頭骨は半円形というか、上縁が丸く下縁がまっすぐ(わずかに凹んでいる)形をしている。このような形の方が、前方で物を咬んだときに吻が変形しにくい。アロサウルスでは後方で咬んだときの頭骨後半部の変形が大きいという。アロサウルスでは咬む力の割に頭蓋が頑丈にできていると昔からいわれているので、それほど新しいことではないが、あらためてアロサウルスの頭骨はなかなかよくできていると思える。そうするとジムマドセニと比べてフラギリスの方が、より変形に強いという意味で頑丈なのだろう。カルカロドントサウルス類も吻の前端は斜めで後頭部が下がっているが、大型化することで全体に頑丈になるのでフラギリスほど頬骨のところが下がらなくてもよいということかもしれない。
参考文献
Fawcett, M. J., Lautenschlager, S., Bestwick, J., & Butler, R. J. (2023). Functional morphology of the Triassic apex predator Saurosuchus galilei (Pseudosuchia: Loricata) and convergence with a post-Triassic theropod dinosaur. The Anatomical Record, 1–17.
https://doi.org/10.1002/ar.25299

コメント ( 0 ) | Trackback ( 0 )
プセウドチャンプサ
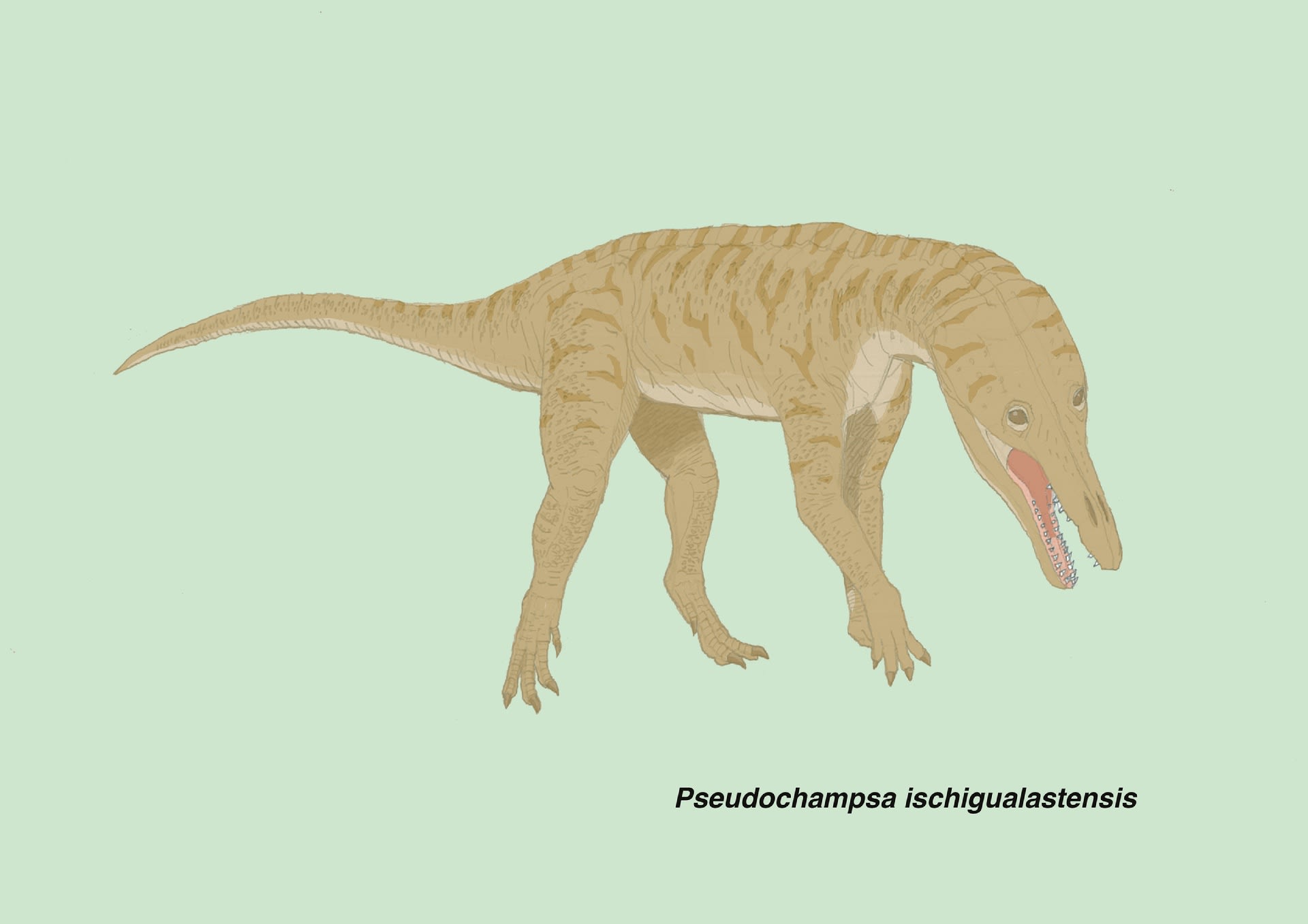
古生物合同展示会は盛況のうちに終了しました。おかげさまで今回、準備したものはほとんどなくなり、特に「三畳紀の爬虫類」シリーズはお手頃価格のカードにしたのがよかったのか、完売しました。お買い上げいただいた方々、ありがとうございました。
やはり、アスファルトヴェナトルとは何か、プロテロチャンプサ類とはどういうものか、口頭で丁寧に説明すると、納得していただけるということで、大変有意義な時間でした。今後もプロテロチャンプサ類をはじめ、「三畳紀の爬虫類」の面白さを広めようと思います。
プロテロチャンプサ類の中でもプセウドチャンプサPseudochampsa ischigualastensisは保存の良い全身の骨格が見つかっており、以前から描きたかったものである。チャナレスクスと近縁で、最初はチャナレスクスと思われたが新属として記載された。ワニのような頭をしたイヌのような動物とされている。頭はワニそっくりだが体は半直立姿勢で、尾も細く水中生活には向いていない。地上でキツネやジャッカルのような役割を果たした捕食者と考えられている。
コメント ( 0 ) | Trackback ( 0 )
アシナシイモリの起源

Copyright 2023 Kligman et al.
現生両生類(カエル、サンショウウオ、アシナシイモリ)は平滑両生類Lissamphibia としてまとめられている。そのうちカエル(無尾類)とサンショウウオ(有尾類)は合わせてバトラキアBatrachiaと呼ばれ、分椎類Temnospondyli のDissorophoidというグループのアンフィバムス科Amphibamidae に近い動物に由来するということがほぼ定説となっている。しかしアシナシイモリ(無足類)の起源については議論があり、分椎類ではない空椎類Lepospondyliに由来するという意見や、分椎類の中でもDissorophoid以外のグループに由来するという意見があり、研究者の間で一致していなかった。
無足類は四肢動物の中でも化石記録が乏しいグループであり、これまではジュラ紀のエオカエシリアEocaecilia が最古の記録であった。エオカエシリアは体が細長いがまだ四肢があり、目も大きかった。しかし歯列や顎の筋肉系など、無足類の特徴を示していた。今回、アリゾナ州の三畳紀後期のChinle formationから、最古の無足類フンクスヴェルミスFuncusvermis が記載された。化石は断片的だが、下顎の骨などが決め手となったらしい。
無足類は二重の歯列をもっている。上顎では上顎骨歯と口蓋歯があり、上顎骨と口蓋骨が癒合して上顎口蓋骨maxillopalatine という骨になっている。下顎では歯骨の代わりに偽歯骨pseudodentary という骨があるが、歯骨とadsymphyseal という骨が癒合しているようだ。歯骨歯とadsymphyseal toothが二重に並んでいる。このadsymphysealという骨は初めて見たので興味深い。
ペルム紀前期の原始的なDissorophoidであるカコプスでは、下顎の歯骨の前端の下顎結合のところに小さな骨がある。これがadsymphysealで、一本の大きな歯が生えている。ペルム紀後期のアンフィバムス類ドレセルペトンDoleserpeton では歯骨歯の内側にいくつかのadsymphyseal toothがある。そしてフンクスヴェルミスでは歯骨歯の内側に多数のadsymphyseal toothが並んでいる。これはジュラ紀のエオカエシリアや後の無足類と同様である。adsymphyseal toothは、カエルやサンショウウオの系統では失われている。
無足類のもう一つの重要な特徴は、顎の筋肉系にある。それはDual Jaw Closure Mechanism(DJCM、二重の顎を閉じる機構)と呼ばれる。他の分椎類では下顎の顎関節の前方に、顎を閉じる筋肉として下顎内転筋musculus adductor mandibulae が付着し、下顎を上に引き上げる。顎関節より後方はあまり突出していない。一方無足類では顎関節より後方に後関節突起retroarticular processという突起が発達しており、これの腹側に舌骨の筋肉が付着している。この舌骨の筋肉が後関節突起を腹側に引くことで、下顎の前方部分が上に回転し、顎を閉じる仕組みである。この二重の筋肉系があることにより、無足類は下顎内転筋を減らすことができ、それは頭骨をコンパクトにするのに役立つとされている。頭を先頭に地中を進む際に、頭骨がコンパクトであることは有利というわけである。エオカエシリアでは後関節突起が発達していたが、フンクスヴェルミスにはなかった。このことからDJCMの獲得は、フンクスヴェルミスとエオカエシリアの間で生じたと考えられる。
現生の無足類では目が退化し眼窩が小さく、眼窩の前方に小さな孔がある。これは化学受容器官である触手tentacle を容れる部分である。ジュラ紀のエオカエシリアでは、眼窩の前方の涙骨のあたりにtentacular fossa という窪みがある。フンクスヴェルミスでは上顎口蓋骨の断片しか見つかっていないが、眼窩の縁が保存されており、眼窩は比較的大きかったことと、tentacular fossaはなかったことがわかった。このことから触手の発達も、フンクスヴェルミスよりは後に起こったと考えられた。
フンクスヴェルミスはいくつかの無足類の形質を示すが、いくつかの形質は欠いていた。また原始的なアンフィバムス類と共有する形質をもっていた。そういうわけで系統解析の結果、フンクスヴェルミスは進化した無足類とアンフィバムス類の中間の位置にきた。またこのことから、現生両生類(平滑両生類)はすべて、分椎類のDissorophoidに由来する可能性が高いと考えられた。
今回は系統学的結論よりも、二重の歯列に感動した。カエルにもサンショウウオにもない歯がアシナシイモリには残っていて、それは古生代の分椎類から受け継いだものである、というところにロマンを感じる。ちなみにシン・ウルトラマンのネロンガだったか、も下顎の歯列が二重になっていたと思うが、アシナシイモリとの収斂でもないだろう。
参考文献
Ben T. Kligman, Bryan M. Gee, Adam D. Marsh, Sterling J. Nesbitt, Matthew E. Smith, William G. Parker & Michelle R. Stocker (2023) Triassic stem caecilian supports dissorophoid origin of living amphibians. Nature Vol 614, 2 February 2023, 102-110.
コメント ( 0 ) | Trackback ( 0 )
シレサウルス (シレサウルス類)
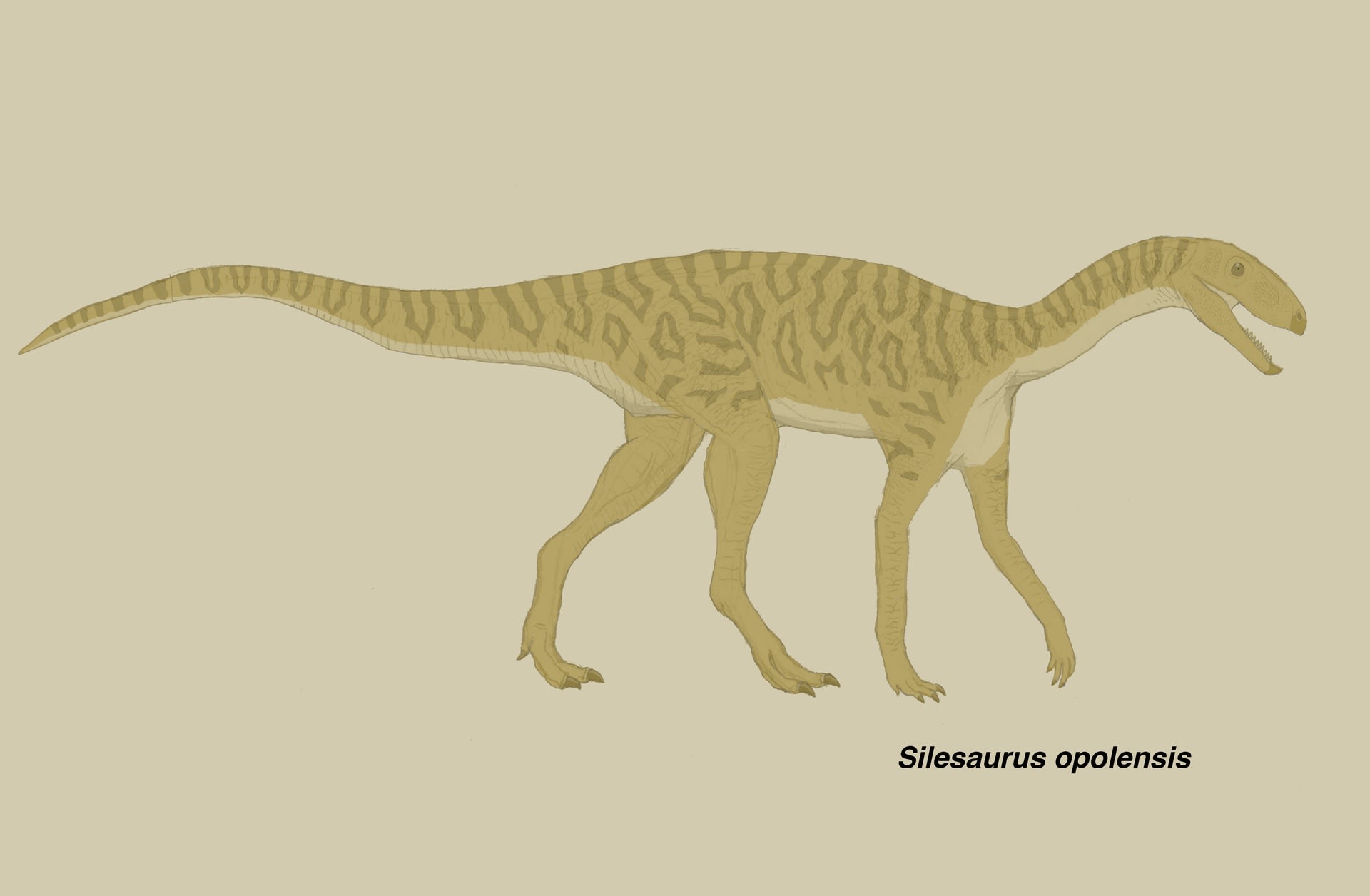
恐竜以外でも肉食にこだわる人が、どういう風の吹き回しかといぶかる向きもあるかもしれない。「三畳紀の爬虫類」カテゴリーでは、「違和感」を描きたいのである。爬虫類のシカのような奇妙な姿は恐竜にはみられないもので、取り上げる意味がある。
シレサウルス類は、一般的には恐竜に最も近縁な恐竜形類であるが、最近のいくつかの研究では鳥盤類の祖先筋である可能性が指摘されている。元々、三畳紀の地層から竜脚形類や獣脚類は発見されているのに、確実な鳥盤類の化石は全く報告されていなかった。鳥盤類の歯とされたものが、実はシレサウルス類の歯であったりした。さらに最も基盤的な鳥盤類であるピサノサウルスが、シレサウルス類に含まれるという研究や、シレサウルス類が鳥盤類に含まれるという研究が報告されている。その場合は単系群であるシレサウルス類が、典型的な鳥盤類(ヘテロドントサウルス以上)と姉妹群をなすという形であった。
その後Müller and Garcia (2020) の系統解析では、シレサウルス類は単系群ではなく多系群となり、典型的な鳥盤類に対して順次外群となった。そしてピサノサウルスはシレサウルス類と典型的鳥盤類をつなぐ位置にきた。また、多くのシレサウルス類は歯の形態から雑食ないし植物食であるが、最も基盤的なレウィスクスLewisuchusは歯の形態から肉食性と考えられている。よって鳥盤類は最も早い時期から雑食ないし植物食を獲得した恐竜であるが、それは祖先的形質ではなく二次的に獲得したものであると思われた。これは竜脚形類も同様である。
シレサウルス類が基盤的な鳥盤類ということになると、従来謎であった三畳紀の鳥盤類が実は見つかっていたことになる。もしそうなら鳥盤類の系統は、三畳紀には四足歩行の植物食動物として十分繁栄していたが、それらは三畳紀末に絶滅し、二足歩行のものだけがジュラ紀まで生存し、再び四足歩行や条件的四足歩行になったということか。シレサウルス類がそのまま生き残ってウシやサイのような姿になっていたら面白かったのだが、それは起こらなかった。大型化という点ではやはり竜脚形類の勢いに圧倒されたのだろうか。
参考文献
Müller RT, Garcia MS. 2020 A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biol. Lett. 16: 20200417. http://dx.doi.org/10.1098/rsbl.2020.0417
コメント ( 0 ) | Trackback ( 0 )
バリナスクス (ノトスクス類、セベコスキア)

解説はChimerasuchusさんの動画が良い。
バリナスクスは新生代の中新世中期にベネズエラ、ペルー、アルゼンチンに生息した最大のセベクス類(またはセベコスキア)で、部分的な頭骨が見つかっている。新生代の初めも島大陸だった南アメリカでは、真獣類が侵入する前、陸上捕食者のニッチをめぐって肉食有袋類、恐鳥類、ワニ類がしのぎを削っていた。ノトスクス類の中でもセベクス類とバウルスクス類は大型の捕食者として進化した。そのうちセベクス類は新生代の前半まで繁栄していたグループである。中でもバリナスクスは中新世中期まで生息していた、陸生ワニ類最後の星である。
バリナスクスの頭骨は約1m 、全長は少なくとも6 m 、体重は1.6-1.7 tと推定されており、新生代を通じて最大の陸上捕食者とされている。完全に哺乳類の時代となってもなお、三畳紀のラウイスクス類の栄光再びとばかりに、頂上捕食者として君臨していたとは、ワニ類のポテンシャルには敬服せざるを得ない。
ネット上のイラストには、セベクスの頭骨そのもののような顔で描かれているものがあるが、不完全ながら頭骨があるのだから、セベクスではなくバリナスクスの頭骨を見て描く方がいいだろう。バリナスクスは少なくとも吻の形がセベクスとは異なる。鼻先が角ばっていて前縁がより垂直に近い。セベクス類やバウルスクス類に共通して頭骨は丈が高く幅が狭く、眼窩は側方を向いている。また歯は側扁したナイフ状で後方に反っていて鋸歯がある。獣脚類と収斂した顔つきである。
アンデス山脈が隆起したなどの影響を受けた気候変化によって、バリナスクスもついに絶滅したらしい。ベネズエラあたりなら低地に移動したり小型化して、なんとか生き残って欲しかった系統である。ワニ類はほぼ熱帯に限られるが、性決定機構が温度に依存することがやはり脆弱性となっているのだろうか。
コメント ( 0 ) | Trackback ( 0 )
ラザナンドロンゴべ (メソエウクロコディリア、ノトスクス類)

(三畳紀ではなくジュラ紀の動物であるが、ワニつながりでここに入れておく。)
恐竜ファンでも知っている人は少ないかもしれないが、ミネラルフェアなどで恐竜の歯を探しているコレクターなら馴染みがあるはずである。私のミネラルフェア・ミネラルショー歴は15年くらいであるが、正体不明の歯化石としてよく見かけるものであった。獣脚類ともいわれるが、違うんじゃないか、と言われていた。結局ワニ類となった。
ラザナンドロンゴベ・サカラヴァエRazanandrongobe sakalavaeは、中期ジュラ紀バトニアンのマダガスカルから産する歯と上顎骨の断片のみの化石として2006年に記載された。特徴的な歯の形状から新属新種として命名されたが、断片的であるために分類学的位置を定めることはできなかった。その後、吻の部分の新たな化石が発見されたことで、多くの情報が得られ、研究が進展した。新たに追加された標本は、ほとんど完全な右の前上顎骨と、左の歯骨の前方部分、そしてホロタイプとは異なる上顎骨の一部である。これらの新しい標本は、2012年にトゥールーズ自然史博物館が個人コレクションから獲得した(寄贈?)とある。
前上顎骨の形から吻は丈が高く、幅広く、尖ってはいなかったことがわかる。前上顎骨には5本の歯があり、それらはほとんど垂直でわずかに内側に曲がっている。前方の歯は断面がU字形で、鋸歯のある稜縁が内側(舌側)にある。骨の外鼻孔は左右が癒合して1つになり、前方を向いている。また前上顎骨/上顎骨境界の側面には、バウルスクス類などにあるような下顎の犬歯状の大きな歯を受け入れる溝はない。歯骨の前方部分には歯のない先端部と8本の大きな歯骨歯があり、そのうち前方の歯(1-3)が最も大きいが、肥大した犬歯状caniniformの歯はない。下顎結合は3番目の歯の位置まで後方に延びている。夾板骨splenialは保存されていないが、歯骨側の関節面から少なくとも下顎結合の長さの20%に寄与していたと考えられた。
ラザナンドロンゴベの歯は非常に太く断面は円形に近い。歯冠には非常に大きな鋸歯があり、ティラノサウルスのような大型獣脚類の最大の鋸歯よりも大きいという。前方の歯は断面がU字形で稜縁が舌側にあり、側方の歯も稜縁が舌側にねじれているため非対称で、中央の断面はサリノン形salinon-shapedで基部は楕円形である。しばしば唇側舌側の幅が前後(近心遠心)の長さよりも大きい。つまりラザナンドロンゴベの歯は多くのジフォスキアやティラノサウルス類よりも肥厚している。
ラザナンドロンゴベの新しい標本は、以下の特徴から確かにワニ形類であり獣脚類ではないことを示している。1)外鼻孔が前方を向き、明らかに癒合していて正中面で左右を隔てる前上顎骨の鼻骨突起がない。2)下顎結合が獣脚類よりも頑丈で、前後に伸びている。3)メソエウクロコディリアと同様に、夾板骨が下顎結合に組み込まれており、下顎の腹内側部分として側面から見えている。4)多くのワニ形類と同様に、上顎骨の骨口蓋がよく発達している。5)歯槽と歯冠が唇側舌側に肥厚し膨張している。これはマダガスカルから知られているアベリサウルス類などの獣脚類とは全く異なる。実際にスピノサウルス類以外の獣脚類ではこのような円錐形の歯はみられないが、ワニ形類には普通にみられる。またスピノサウルス類の前上顎骨は、外鼻孔が後退しているなど多くの点でラザナンドロンゴベとは異なる。
ラザナンドロンゴベでは外鼻孔が前方を向いている。この形質は多くのワニ形類にみられる。バウルスクス科Baurusuchidaeでも外鼻孔が前方を向いているが、前上顎骨と鼻骨からなる中隔で仕切られている。セベクス科Sebecidaeでは外鼻孔は仕切られており、また背方を向いている。スファゲサウルス科Sphagesauridaeはラザナンドロンゴベと同様に、左右の外鼻孔が癒合し、前方を向いている。
まだまだ断片的ではあるが系統解析の結果、ラザナンドロンゴベは、ワニ形類、メソエウクロコディリア、ノトスクス類となった。知られる限り最古のノトスクス類であるという。ノトスクス類は主に白亜紀のゴンドワナ地域で繁栄した、植物食のキメラスクス、雑食のシモスクスから肉食のバウルスクスまで含む、非常に多様な陸生ワニ類のグループである。ノトスクス類の中でも、ラザナンドロンゴベはジフォスキアZiphosuchiaの中でセベコスキアSebecosuchia(セベクス科+バウルスクス科)と近い位置にきた。よってセベクスやバウルスクスのような肉食性の種類に近縁だが、それらのもつ犬歯状の大きな歯はないらしい。
ラザナンドロンゴベの頭骨は復元すると1m以上になり、バリナスクスよりもやや大きい最大のノトスクス類となった。これより大型のワニはサルコスクスやプルスサウルスなど、ずっと時代の新しい半水生の種類だけのようである。
参考文献
Dal Sasso et al. (2017), Razanandrongobe sakalavae, a gigantic mesoeucrocodylian from the Middle Jurassic of Madagascar, is the oldest known notosuchian. PeerJ 5:e3481; DOI 10.7717/peerj.3481
コメント ( 0 ) | Trackback ( 0 )
スクレロモクルスの復活
何が復活なのかというと、翼竜の祖先に近い動物として、復活した。
スクレロモクルスは、100年以上前にスコットランドで三畳紀後期の地層から発見された20 cmほどの小型の爬虫類で、後肢が長く二足歩行に見えることから、古くから恐竜あるいは翼竜の祖先と想像されてきた。スクレロモクルスは、若い人よりもおっさん世代の恐竜ファンの方が馴染み深いかもしれない。昭和の恐竜本では、よく木に登ったり地上を走ったりしていた動物である。7個体の骨格が知られているが、非常に小型であるうえにつぶれており、詳細な解剖学的特徴を定めるのが困難であった。その復元姿勢や行動様式については、二足歩行か四足歩行か、這う姿勢か直立姿勢か、地上性か樹上性かなど多くの議論があった。つい最近は、走行性ではなくカエルのように跳躍する主竜形類とする研究が提唱されていた。つまり恐竜や翼竜とはかけ離れた系統上の位置になっていた。それが、今月になって翼竜形類つまりラゲルペトン類に近い動物として復帰したのである。
スクレロモクルスの化石は岩にへばりついた印象化石のようなもので、骨の立体的な形態を論じるには多くの困難があった。これまでの研究は表面から2次元的に観察したものだった。Foffa et al. (2022) は模式標本をマイクロCTでスキャンし、初めて全身骨格の3次元再構築を行った。その際、まだ母岩に埋もれていた足の指骨、肋骨、尾椎などが新たに発見され、それらを元にこれまで知られていなかった形態学的特徴が得られた。例えば肋骨は、従来の想定よりも3~4倍も長いことがわかり、胴はカエルのように扁平ではなく、丈が高いことがわかった。頭骨も上下につぶれているが、復元するとかなり丈が高いと考えられた。完全な足は初めて得られたものであり、第4指が最も長くラゲルペトン類と似ていた。
系統解析の結果、スクレロモクルスは少なくとも8つの共有派生形質で翼竜形類(ラゲルペトン類+翼竜類)に属することが支持された。上顎骨の上行突起の基部が凹型、第1中手骨が細長い、恥骨が大腿骨の50%より短い、大腿骨頭がカギ形hook-shaped、第5中足骨が短い、などである。足根部の構造はマイクロCTを用いてもはっきりせず、ワニ型とも恐竜型とも解釈できる。ほとんどの解析結果では、スクレロモクルスは基盤的なラゲルペトン類に含まれた。足根部がワニ型とスコアした場合でも、スクレロモクルスは翼竜形類全体と姉妹群となった。
過去の研究で四足歩行、這う姿勢、カエルのような跳躍性とする説の多くは、不完全なデータ(短い肋骨、扁平な胴と頭骨)やあいまいな解釈(足根部)に基づいたものであるという。またスクレロモクルスは、樹上性の動物がもつ形質(曲がったカギ爪や長い指など)は示さない。スクレロモクルスは後肢の長さに対して腰帯が小さいが、カエルやトビネズミのような跳躍性の動物は、小型であっても大きな腰帯と筋付着部をもっている。
ただし前肢と後肢の長さの比率は四足歩行と二足歩行の中間であるなど、はっきりしない点もある。著者らはスクレロモクルスは、指行性、地上性、走行性であり、少なくとも条件的二足歩行であっただろうとしている。
論文には立派な復元画があるが、羽毛も生えて小鳥のような可憐な姿になっている。2020年に扁平なカエル形の動物だったものが、2022年にここまで劇的に変わるとは、話が違うじゃないか、という感じである。古生物の復元がいかに難しいか、というありがちな文言を言わざるを得ない。姿勢に関係する大腿骨頭や膝関節のあたりも、今回のマイクロCTで情報が得られた部分らしいので、やはり3次元の威力ということか。
参考文献
Foffa, D., Dunne, E.M., Nesbitt, S.J. et al. Scleromochlus and the early evolution of Pterosauromorpha. Nature (2022). https://doi.org/10.1038/s41586-022-05284-x
コメント ( 0 ) | Trackback ( 0 )
スモック(スモク、スモーク)

昨年の三畳紀の爬虫類のときの、ボツ作品。
解説はDr. Polaris の動画、early archosaurians だったか、を見るのがわかりやすい。
三畳紀後期のポーランドの頂上捕食者で、これのものとされる糞化石の分析から、サイほどもある大型単弓類リソウィツィアや迷歯類などを捕食していたことがわかった。つまり、ティラノサウルスのように獲物を骨ごと噛み砕いて食べていた。
最初の記載論文では偽鰐類Pseudosuchia でもアヴェメタターサリアAvematatarsalia でもない、基盤的な主竜類とされたが、ラウイスクス類に過ぎないとの意見もあるらしい。ワニ系でも恐竜系でもない最強動物として、エキゾチックな魅力があると思ったが、ラウイスクス類とすればあまり面白くない。首をS字状に持ち上げると恐竜的になり、水平にするとワニ的になるが、これを描いた時は微妙に折衷案にしたのか。
コメント ( 0 ) | Trackback ( 0 )
ラゲルペトン類 (アヴェメタターサリア、翼竜形類)

最新の研究で、翼竜の祖先に最も近いとされる動物。
ラゲルペトン類は、小型から中型(通常1m以内)のほっそりした走行性の爬虫類で、三畳紀中期から後期の南アメリカ、北アメリカ、マダガスカルから知られている。過去にはラゲルペトン類については、脊椎、後肢、わずかな頭の骨しか知られておらず、恐竜形類とされていた。近年、ラゲルペトン類と初期の翼竜の化石が発見され、詳細に研究できるようになったため、Ezcurra et al. (2020) はこれらの標本をマイクロCTなどを駆使して解析し、ラゲルペトン類と翼竜類が姉妹群をなすという結果を報告している。これらの結果はラゲルペトン、イクサレルペトン、コンゴナフォン、ドロモメロンなどという聞きなれないラゲルペトン類の全身の骨格に基づいている。論文の全身復元骨格も、1種類の全身骨格ではなくて、ラゲルペトン、イクサレルペトン、ドロモメロンを合成したものである。
ラゲルペトン類と初期の翼竜類とは、例えば以下のような形質を共有している。
1)上顎骨の前方突起の大部分が、外鼻孔に面している。
2)歯骨の前端には歯がなく、先細りに尖った形をしている。
3)歯骨の前方部分が腹側に曲がっている。
4)歯骨の前方の歯が(マシアカサウルスのように)前傾している。
5)下顎の夾板骨splenialがないか、退化的である。
6)歯骨の歯の数が多い(20以上)
7)歯列の後方の歯冠には3つの咬頭がある。
8)歯間板がない。
9)よく発達し後側方に尖った小脳片葉をもつ。
10)内耳の三半規管の部分は前後の長さより高さが大きく、前方半規管は後方半規管よりもかなり長いアーチをなす。
11)前肢の前腕は上腕骨よりも長い。
12)中手骨は比較的長く伸びている。
13)腰帯の恥骨と坐骨の結合が、腹側縁まで伸びている。
14)大腿骨頭がカギ形(hook-shaped)である。
15)距骨と踵骨が癒合(co-ossified)している。
これらの中で、おっと思ったのは歯の形状である。ラゲルペトンやイクサレルペトンでは歯列の後方の歯が3つの咬頭をもつ。複数の咬頭をもつ歯は主竜形類には珍しいが、初期の翼竜ラエティコダクティルス、カルニアダクティルス、エウディモルフォドンなどにはみられる。こういう飛行と関係ない部分に共通点があるとそれらしい気がする。
系統解析の結果、ラゲルペトン類が翼竜類と姉妹群をなすことが強く支持された。両者を含むクレード、翼竜形類Pterosauromorpha は、少なくとも33の共有派生形質で支持される。これらのうちいくつかの形質は、翼竜形類に固有であるという。内耳の三半規管の高さと長さの比率が0.9より大きい、夾板骨が退縮または欠損している、恥骨と坐骨の結合部pubo-ischial plateが腹側に伸びている、大腿骨頭がカギ形などである。
最も近縁といっても、ラゲルペトン類と翼竜類ではまだまだ多くの形態学的ギャップがある。ラゲルペトン類には、翼竜のような上腕骨の三角筋稜の拡大や第4指の拡大はみられない。第4指が少し伸びていたりすると感動するのだが、そうではない。これはやはり、樹上性とか滑空性という過程があるのだろう。地上に留まったのがラゲルペトン類で、樹上性から滑空に至ったのが翼竜類ということか。第4指が伸びるのは、皮膜で滑空することを前提とした変化のような気はする。ドロモメロンでは手の末節骨の曲がりが強いことから、木登りや獲物の捕獲などに用いた可能性があるといっている。
参考文献
Ezcurra, M.D., Nesbitt, S.J., Bronzati, M. et al. (2020) Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature 588, 445–449. https://doi.org/10.1038/s41586-020-3011-4
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ |