獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年9月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
スミロドンはガオーと吠えたかニャーと鳴いたか

Copyright 2023 Deutsch et al. 上がトラ、下がカラカルの舌骨
動物好きの方なら聞いたことがあると思うが、ネコ科の動物にはガオーと吠えるものと、ニャーと鳴くものがいる。イエネコを含むヤマネコの仲間はニャーと鳴き、ノドをゴロゴロ鳴らす(pur)ことができる。一方ライオンやトラのようなヒョウ属Pantheraの大型ネコ類は、ガオーと吠える(roar)が、ノドをゴロゴロ鳴らすことはできない。これは舌骨の構造が違うためである。舌骨はいくつかの骨からなるが、小型ネコ類では上舌骨epihyoidがあるが大型ネコ類では上舌骨がなく靭帯になっているためといわれている。体は大きくてもピューマはニャーと鳴き、ノドをゴロゴロ鳴らす。またチーターがニャーと鳴く動画はYouTubeで見ることができる。
ロサンゼルスのランチョ・ラ・ブレアRancho la Brea のタールピットからは、多数の肉食獣の化石が見つかっており、サーベルタイガーの代表であるスミロドンSmilodon fatalis の舌骨も多数見つかっている。そこでスミロドンがどのような鳴き声で鳴いたかという研究がされてきた。2018年の研究ではスミロドンでは上舌骨が発見されないことから、ヒョウ属のように吠えただろうと推定された。しかしこの時は舌骨の各骨の形態について、詳細な解析はされていなかった。そもそもスミロドンはネコ科の中で初期に分岐したマカイロドゥス亜科に属しており、トラに近いわけではない。イエネコとトラの方が互いに近縁で、スミロドンからは離れているのである。
そこでDeutsch et al. (2023) は、現生のライオン、トラ、ヒョウ、ジャガー(以上ヒョウ属)、ピューマ、チーター、カラカル、サーバル、オセロットと、タールピットの化石種であるスミロドン、アメリカライオンPanthera atrox の舌骨の各骨について、定量形態学的に比較した。まずスミロドンの舌骨は現生種よりも全体に大きく太いことから、より低音で鳴いたと考えられた。次に各骨の形態を多変量解析した結果、系統的にライオンに近いアメリカライオンは確かにヒョウ属に近い形態といえるのに対して、スミロドンの舌骨は多くの点で小型ネコ類と似ていることがわかった。これらのことから、スミロドンはトラのように吠えたのではなく、低音でニャーと鳴いた可能性があるという。
参考文献
Deutsch, A. R., Brian Langerhans, R., Flores, D., & Hartstone‐Rose, A. (2023). The roar of Rancho La Brea? Comparative anatomy of modern and fossil felid hyoid bones. Journal of Morphology, 284, e21627. https://doi.org/10.1002/jmor.21627
コメント ( 0 ) | Trackback ( 0 )
プシッタコサウルスとレペノマムスの格闘化石
中国遼寧省でまた、恐ろしく保存のよい化石が発見され、恐竜と哺乳類の関係の見方に一石を投じるものとしてニュースになっている。
発見された化石は2頭の動物が密接に絡み合ったもので、1頭は小型のケラトプス類プシッタコサウルスPsittacosaurus lujiatunensis(120cm)、もう1頭はより小型の哺乳類レペノマムスRepenomamus robustus (47cm)である。
プシッタコサウルスの骨格はほとんど完全で、うつ伏せに横たわっており、頭と尾は左に曲げている。大腿骨の周長などから体重は10.6 kgと推定された。レペノマムスの骨格もほとんど完全で、大きく体を曲げており、恐竜の体の左側の上に乗っていた。下顎や歯の特徴は埋まっていてわからないが、比較的小型であることと矢状稜や頬骨弓の発達が弱いことからレペノマムス・ロブストゥスであり、さらに大きなレペノマムス・ギガンティクスではない。推定体重は3.4 kgで、前頭骨の縫合はそれほど顕著でないが長骨の骨端が閉じていることから亜成体と考えられた。レペノマムスの左手はプシッタコサウルスの下顎をつかんでおり、左足はプシッタコサウルスの膝関節に挟まれて、脛をつかんでいる。またレペノマムスの顎はプシッタコサウルスの前方の肋骨2本を咬んでいた。(写真の説明ではなぜか前腕を咬んでいるように書いてあるが、本文では一貫して肋骨になっている。直し忘れか)
この化石は発見者から博物館に寄贈されたものであるが、2種類の動物が密接に絡み合っていることから、過去の遼寧省にあったような贋作化石ではなく、本物であると考えられた。著者らは本物である確信を得るため、まだ母岩に埋まっていた下顎をクリーニングしたが、やはり肋骨を咬むように位置していた。
また化石化の過程で川の流れによって単に2つの死骸が寄せ集められたものでもない。骨格がほとんど完全に保存されていることから、これは明らかに長距離を流されたものではなく、死亡した場所で火砕流などに埋もれたものと考えられた。
2種の動物が相互作用したとしても、いくつかの可能性があり、例えばプシッタコサウルスの死体をレペノマムスが食べていた(屍肉食)のかもしれない。この仮説はプシッタコサウルスの方がレペノマムスよりもずっと大きいこと、生きた動物の肋骨を咬むのは困難であるように思われることと整合する。しかし著者らは、プシッタコサウルスの骨に咬み跡がないこと、これらの動物が複雑に絡み合っていること、レペノマムスがプシッタコサウルスの上に乗っていることなどから、実際に捕食の過程を表しているという説を支持している。なるほど死体を漁るだけだったら、下顎をつかんだり足が恐竜の膝に挟まれたり、こんなアクロバティックな姿勢になることはないように思える。
プシッタコサウルスの方がずっと大きいことについては、著者らは捕食者と被捕食者のサイズ関係について多くのデータベースを検討した結果、ありえないことではないと言っている。現生種でも例えばクズリは、自分より数倍も大きいヘラジカやトナカイを襲うことが知られている。イタチ科で最小のイイズナも、自分より大きいライチョウやノウサギを襲うことがある。そういえばレペノマムスはよくタスマニアデビルのようながっしりした体形といわれるが、クズリのように気が強いのかもしれない。
肋骨を咬んでいることについては、獲物が生きたままの状態で食べ始めることは、リカオンやブチハイエナにみられるといっている。最初に捕食者からの攻撃を何度も受けた獲物は、体力の消耗とショックにより倒れてしまうことがある。急所であるノド付近を食い破り、さらに胸部をかじっている状態なのだろうか。そうだとすれば凄まじい攻撃の瞬間を見ていることになる。プシッタコサウルスの上にレペノマムスが乗っているのは、弱った獲物を制圧しているとも見えるという。
マイケル・スクレプニクさんの素晴らしい復元画が載っている。足を恐竜の膝に絡めてほしかったな。
参考文献
Gang Han, Jordan C. Mallon, Aaron J. Lussier, Xiao‑Chun Wu, Robert Mitchell & Ling‑Ji Li (2023) An extraordinary fossil captures the struggle for existence during the Mesozoic.
Scientific Reports (2023) 13:11221 https://doi.org/10.1038/s41598-023-37545-8
コメント ( 0 ) | Trackback ( 0 )
ゴルゴノプスの胴体

Copyright 2023 Bendel et al.
ゴルゴノプスのほとんど完全な胴体、素晴らしいですね。
前肢の骨は太くがっしりしているのに対して、後肢の骨は細長いことから、ゴルゴノプスは待ち伏せ急襲型のハンターで、短距離だけ獲物を追跡し、力強い前肢で倒してから犬歯で仕留めたと考えられる。推定体重は98 kgで雌ライオンと同じくらいという。
ゴルゴノプス類は大体同じような体形をしていて、胴体の骨だけで種類を区別するほどの特徴をあげるのは難しい。しかし細かい差異がないわけではない。
胴体の骨格で最も特徴的なのはイノストランケビアで、肩甲骨が幅広いので他のゴルゴノプス類とは区別できる。また前肢の橈骨と後肢の脛骨が太いという。まあがっしりした体格ということでしょう。ロシア産の種類でもサウロクトヌスなどの肩甲骨は、若干の特徴はあるがアフリカ産のゴルゴノプス類と似たプロポーションのようである。
アフリカ産のゴルゴノプス類の中でも肩甲骨の形状には変異がある。また今回の発見により明らかになった点として、ゴルゴノプスでは手根部の橈側骨radialeが三角形である。末節骨はゴルゴノプスでは太く短いが、スキムノグナトゥスでは長く鋭いことから、捕食の際の使い方に違いがあった可能性がある。
また腰帯をみると、リカエノプスなどでは恥骨と座骨の間がくびれているが、ゴルゴノプスではほとんどくびれていないのが特徴である。
参考文献
Bendel E-M, Kammerer CF, Smith RMH, Fröbisch J. 2023. The postcranial anatomy of Gorgonops torvus (Synapsida, Gorgonopsia) from the late Permian of South Africa. PeerJ 11:e15378 DOI 10.7717/peerj.15378
コメント ( 0 ) | Trackback ( 0 )
有胎盤哺乳類の脳化の様相:まず体が大きくなり、それから脳が発達した

バリラムブダ(汎歯類)

フェナコドゥス(か節類)

パトリオフェリス(オキシアエナ類、始新世の原始的な肉食獣)
写真は全てwikipedia (en.wikipedia org)
哺乳類は、脊椎動物の中で、最も体の割に大きな脳をもつ動物である。哺乳類の脳の進化については、中生代の間にもいくつかの段階で発達がみられるが、それは現代の哺乳類に比べるとはるかに原始的な脳であった。哺乳類の脳が劇的に発達するのは新生代に入ってからであるという。科学番組などでは恐竜時代に、哺乳類は小型で夜行性の生活をしていたために、聴覚や空間認識など情報処理能が発達したことが紹介されていた。それがそのまま発展したのかというと、そう単純でもないらしい。
白亜紀末に恐竜が絶滅した後、生態系の空白のニッチを埋めるように哺乳類が多種多様に放散した。その後も哺乳類の進化が続いて今日に至るわけだが、その過程で哺乳類の脳がどのように発達したのかについては、詳細が知られていなかったという。
Bertrand et al. (2022) は、中生代から恐竜絶滅後の古第三紀暁新世、始新世にかけて、多数の有胎盤類の脳函をCTスキャンし、脳容量endocranial volume、推定体重、脳化指数PEQ(体の大きさに対する相対的な脳の大きさ)を計算した。最近ニューメキシコ州で発見された、新しい暁新世の哺乳類の脳函も含まれている。その多数のデータを中生代哺乳類、暁新世の哺乳類、始新世の基盤的な哺乳類、始新世の進化的哺乳類に分けてグラフで示している。また脳全体の容量だけでなく、脳の各領域(嗅球など)の容積を算出して各系統で比較した。
従来から、暁新世の哺乳類の脳は中生代哺乳類とあまり変わっておらず、そのまま拡大したバージョンであるという説は提唱されていた。しかしBertrand et al.の結果は少し異なっていた。体のサイズは、中生代に比べて、古第三紀ではみな非常に大きくなっているが、相対的な脳の大きさは、暁新世では中生代よりもむしろ低下しており、始新世になると増加している。そして始新世の中では、原始的な系統に比べて、進化的な系統では明らかに相対的な脳の大きさが増加していた。
脳の各領域についてみると、始新世の進化的な系統では、暁新世や始新世の原始的な系統に比べて変化していた。相対的に嗅球が小さくなり、錐体小葉petrosal lobuleや新皮質が大きくなっていた。錐体小葉は平衡覚、視覚、眼球運動、頸の動きなどに関与する部位である。また新皮質は情報の統合にあずかる。つまり暁新世までは嗅覚に依存した原始的な脳であったが、始新世の一部の系統でその他の感覚や情報処理が大きく発達したということになる。
暁新世における相対的な脳サイズの低下には、“か節類”(ex.フェナコドゥス)、汎歯類(バリラムブダ)、紐歯類(スティリノドン)などの原始的な系統で体のサイズが増大したことが関与している。一方始新世になると、特に進化的な系統で相対的な脳サイズが増加した。例えば偶蹄類(クジラ類を含む)、奇蹄類、食肉類、真霊長類などである。また暁新世では、雑食及び肉食の種類と草食の種類で相対的な脳サイズに有意の差がなかったが、始新世になると雑食及び肉食は、草食を凌駕するようになった。
恐竜絶滅後の哺乳類の放散においては、脳のサイズよりも体のサイズの増加が特徴的であるともいえる。体が大きくなるにつれて脳も大きくなるが、構造的には変化がなかった。独立して体が大きくなったことで、相対的な脳サイズには変異が大きくなった。暁新世の哺乳類にとっては、相対的に大きな脳は必要でなかったかもしれないという。
始新世になると、空いていたニッチが飽和し、種間の競合が起きるようになった。相対的な脳サイズが大きく、また感覚や情報処理、社会行動などの能力がより優れた種類が有利なように自然選択が働くようになったという趣旨である。
感想であるが、確かにバリラムブダのような暁新世の草食哺乳類をみると、頭が小さく、とりあえず体を大きくしたようなバランスが感じられる。正確には植物食の単弓類に近い体形なのか。このままさらに大型化して恐竜的な存在になってくれてもよかったのだが、そこまで生物資源が豊かではなかったか、時間がなかったのだろう。始新世には脳が発達した新しいグループが出現して、競合にさらされ、後に取って代わられることになった。
肉食獣の方は外見ではわからないが、脳のサイズに差があるのだろう。ヒエノドン類と食肉類ではPEQに差があることがグラフに出ている。ただし、ヒエノドン類でも一部の種類は食肉類と遜色ないくらいの脳サイズだったようだ。
そうすると現代型の哺乳類の脳が発達したのは、恐竜におびえて夜の世界に暮らしていたからというよりも、始新世に哺乳類同士の競争が激しくなったからということになる。ジュラ紀や白亜紀の恐竜の間でも熾烈な競合はあっただろうが、脳サイズが増大する方向に進化したようにはあまり見えない。
真獣類全体の脳の初期進化を扱ったすごい研究だと思ったら、獣脚類の研究者でもあるエジンバラ大学のBrusatte博士のラボだった。ただアフリカ獣類や異節類のデータがほとんどないのは、暁新世や始新世の化石が乏しいのだろう。
参考文献
Bertrand et al. (2022) Brawn before brains in placental mammals after the end-Cretaceous extinction. Science 376, 80–85, 1 April 2022
コメント ( 0 ) | Trackback ( 0 )
単孔類の祖先も乳を吸っていた
Scienceの News にGretchen Vogel氏が寄稿しています。ハリモグラの赤ちゃんの写真が意外とかわいいので、読む気になりました。
哺乳類の赤ちゃんは母乳を飲む。乳を吸うことは哺乳類の特徴とされている。しかしカモノハシやハリモグラのような単孔類は、乳を吸わない。単孔類にはまとまった乳頭がなく、新生児は母親の皮膚に分布する乳腺からしみ出るミルクをなめとっている。単孔類が哺乳類の祖先から分岐したのは1.9億年前と考えられているので、乳を吸う能力はその後になって進化したと考えられてきた。
ところが、ハーバード大学のCromptonらのグループの最新研究により、単孔類の祖先も乳を吸う能力を持っており、単孔類はそれを二次的に失った可能性が出てきた。
乳を吸う能力の獲得には、口蓋や舌などに関係する骨や筋肉など、かなり複雑な解剖学的変化が必要である。その一つ、tensor veli palatiniという筋肉は、軟口蓋を緊張させることで舌との間に密閉空間を作り、ミルクを吸引するのに役立っている。
Cromptonらは有袋類オポッサム、単孔類カモノハシ、爬虫類オオトカゲの口腔部分を詳細に比較した。さらに三畳紀のキノドン類トリナクソドン(2.5億年前)とブラジリテリウム(2.2億年前)の口腔部分とも比較した。その結果、トカゲとトリナクソドンにはtensor veli palatiniはなかったが、ブラジリテリウムの口蓋の骨の形態や筋付着部から、ブラジリテリウムには原始的なtensor veli palatiniがあったと考えられた。つまりブラジリテリウムは、口腔内にミルクを吸引することができたかもしれない。
哺乳類の共通祖先が現れたのは2億年前と推定され、その後単孔類が分岐したとされるので、単孔類は二次的にtensor veli palatiniを失ったと推測される。つまり乳を吸う能力はまさに哺乳類の祖先がもっていた一連の形質の一つであったということになる。
またカモノハシの口腔部分を研究した結果、単孔類は特殊な摂食方式のために乳を吸う能力を失った可能性も考えられた。カモノハシは、顎を動かすことで口蓋と舌のパッドで甲殻類の硬い殻を挽きつぶす。
これは、単孔類の分岐年代がさらに古かったり、ブラジリテリウムだけが特殊な種類だったりするとまた変わってくるのでしょう。また口蓋だけでなく、化石には残らない乳腺や乳頭の進化とも関連しているはずなので、そこが難しいような気がする。
参考文献
Gretchen Vogel (2018) Echidnas don't suck—but their ancestors did.
Science 20 Jul 2018: Vol. 361, Issue 6399, pp. 213 DOI: 10.1126/science.361.6399.213
最初に掲載された時のタイトルは
Got milk? Even the first mammals knew how to suckle
コメント ( 0 ) | Trackback ( 0 )
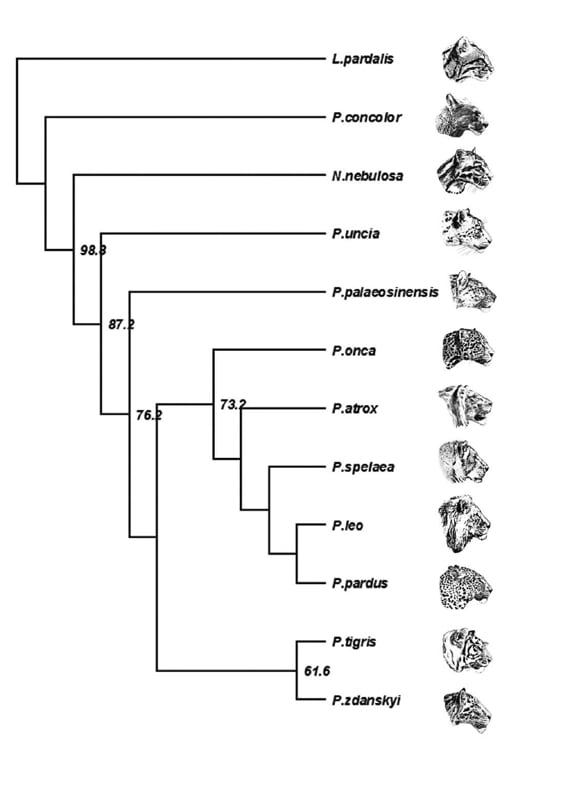
パンテラ・ズダンスキィ(トラの祖先)

Copyright 2011 Mazak et al.
獣脚類でも恐竜でもないが、肉食性の哺乳類、特にネコ科も好きなので。
大型ネコ類であるヒョウ亜科は、現生種の他に、主に更新世中期から後期のいくつかの化石種を含む。また現生種と考えられるすべての化石は更新世のものである。
更新世前期、または鮮新世/更新世境界ごろのパンテラ・パレオシネンシスという動物が、トラの系統の最古のメンバーと考えられてきた。しかし最近の分岐学的、定量形態学的解析によると、パンテラ・パレオシネンシスはトラに近縁ではなく、ヒョウ亜科の中でより基盤的な位置にくるという。確かにトラと考えられる最古の化石は、更新世前期 (Calabrian) の中国・藍田から見つかった上顎と下顎の断片であるが、ある程度完全な頭骨はすべて更新世中期か後期のものである。現生のトラの亜種は形態学的、遺伝学的な差異を示すが、ずっと遅く10万年ほど前に分岐したと推定されている。
2004年に中国甘粛省のLongdanで、哺乳類化石を含む更新世前期の層が発見され、古地磁気学的データから255-216万年前と推定された。これは伝統的には鮮新世の末期であるGelasianに相当するが、Gelasianは最近では更新世初期とされている。化石の中にはヒョウ類の吻部があり、最初は大きさからパンテラ・パレオシネンシスと思われた。しかしその後、完全で保存の良い頭骨が発見されたことにより、これらはパンテラ・パレオシネンシスではなく、ジャガーほどの大きさのヒョウ類の新種であり、パレオシネンシスよりも形態学的にずっとトラに似ていることがわかってきた。
パンテラ・ズダンスキィの分類学的特徴は、以下のようである。ジャガーくらいの大きさのヒョウ類で、頑丈な頭骨をもつ;よく発達した頭部の稜;大きく頑丈な犬歯;頭骨長に対して長い鼻骨;鼻骨が上顎骨-前頭骨の縫合よりも後方に延びている;ハート形の外鼻孔;腹側の輪郭がまっすぐで頑丈な下顎;相対的に大きな裂肉歯と全般に大きな歯;上の裂肉歯P4に顕著なectoparastyleとよく発達したprotoconeがある;下の裂肉歯M1にはっきりしたtalonidと、短く低いparaconid と protoconidがある(以下、歯の咬頭の名称がわからないので略)。
どこがトラと似ているのだろうか。パンテラ・ズダンスキィではトラと同様に上顎の犬歯がよく発達して大きく、その歯冠の高さが頭骨の長さ(condylobasal skull length, CBL)の23.7%に達する。これは現生のトラ(0.166–0.230)よりもむしろ大きく、ジャガー(0.160–0.206)、ヒョウ(0.132–0.202)、ライオン(0.141–0.185)、パレオシネンシス(0.200)よりもずっと大きい。もう一つのトラに特徴的な形質は、頭骨長に対して鼻骨が長いことである。パンテラ・ズダンスキィでは鼻骨の長さはCBL の34.5%であり、これは現生のトラ(0.333–0.417)としては低い方の範囲に入るが、ジャガー(0.275–0.346)、ヒョウ(0.296–0.347)、ライオン(0.287–0.357)の上限にあたり、P. atrox やP. spelaeaよりも大きい。パンテラ・ズダンスキィでは、鼻骨が上顎骨-前頭骨の縫合よりも後方に延びているが、これもトラの特徴である。パレオシネンシスでは鼻骨と上顎骨-前頭骨の縫合は同じくらいの位置にある。さらに頬骨弓が太い点もトラと似ている。
下顎はがっしりしており、腹側の輪郭がまっすぐである点でトラと似ている。これまで、原始的なトラは現生のトラと比べて比較的小さい裂肉歯をもっており、大きい裂肉歯をもつトラは更新世後期になって初めて現れたと考えられていたが、パンテラ・ズダンスキィはそうではないことを示している。パンテラ・ズダンスキィの上の裂肉歯P4の長さはCBL の13.4%で、これはトラ、ジャガー、ヒョウ、ライオンなどの範囲の大きい方に入る。下の裂肉歯M1は下顎長の14.7%で、ヒョウ属の他の種よりも大きい。興味深いことに、パンテラ・ズダンスキィの下顎の歯列は、非常に短く低いparaconid と protoconidをもつ点で、他のヒョウ属と比べてユニークである。
パンテラ・ズダンスキィの標本が完全であることで、これまでよりも総合的にトラの形態進化について推測できるようになった。トラはもともとジャガーほどの大きさの、頑丈な頭骨をもつ動物で、トラ的な頭骨と上顎歯列は進化の初期から獲得していた一方で、下顎と下顎歯列はまだ原始的で、その後の進化で急速に変化したと考えられる。同じようなモザイク的進化はチーターでも知られており、原始的なチーターAcinonyx kurteniはチーター的な頭骨と原始的な歯列をもっていたことから、このようなパターンはネコ科の進化に共通しているかもしれない、としている。
参考文献
Mazak JH, Christiansen P, Kitchener AC (2011) Oldest Known Pantherine Skull and Evolution of the Tiger. PLoS ONE 6(10): e25483. Doi: 10.1371/journal.pone.0025483
Copyright 2011 Mazak et al.
コメント ( 0 ) | Trackback ( 0 )