獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年9月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||
| 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 21 | 22 | 23 | 24 | 25 | 26 | 27 | ||
| 28 | 29 | 30 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
マイプ対ピクノネモサウルス

最大のメガラプトル類と最大のアベリサウルス類、夢の競演。
どちらも想像というところが痛いが、夢だから。
社交ダンスしているのではなく、メガラプトル類としては背後から抱きつくようにして首の血管を切り裂く、必殺技を仕掛けようとしている。でも優しくエスコートしているようにも見えてきた。
コメント ( 0 ) | Trackback ( 0 )
プウィアンヴェナトルの森

プウィアンヴェナトルは、前期白亜紀バレミアン(Sao Khua Formation)にタイ東北部に生息した最も原始的なメガラプトル類である。推定全長6 m で骨格はアウストラロヴェナトルと似ている。
プウィアンヴェナトルは嘆いた。腹が減った。そろそろ虫でもカエルでも食べないと。昨日はせっかく手頃な竜脚類の幼体を見つけたのに、カルカロ族の若者に目の前で横取りされた。このあたりには昔はメトリア族がいたが、最近は北西の乾燥地帯からやってきたカルカロ族が幅をきかせている。カルカロ族は竜脚類を狩るのにたけており、卵、孵化したての幼体からかなり大きい亜成体、老齢の個体まで無駄なく食物資源として活用する。カルカロ族の幼体は小型の獲物を消費するので、彼らが分布する地域では我々中型のハンターは商売あがったりで、数が減ってきていた。
昆虫くらいはそこらにいるが、栄養価が低くて割に合わない。常に大集団で暮らすシロアリのような昆虫が、一定の場所に巣を作っていればいいのだが。昆虫食に適応した者たちは体のサイズが小型化し、発達した聴覚と視覚で虫を探索する夜行性のハンターになっていった。また植物食に走った者たちは、歯がギザギザになるか退化傾向を示し、長い首と大きなカギ爪のある手で枝を手繰り寄せて、もしゃもしゃと食べている。やはり大量に食べる必要があり、消化に時間がかかるのででっぷりメタボな腹部をしている。ああはなりたくないなあ。しかしあの巨大なカギ爪は、強力な武器にもなりそうだ。
プウィアンヴェナトルは肉が食べたかった。大型の獲物を狙うなら、カルカロ族のように大きな顎とナイフのような歯で肉片を切り取るのがいいだろう。彼らは前足はあまり使わないので、小さくなっていた。しかし自由度の高い前足で、小型の獲物を捕らえるのは三畳紀以来、獣脚類の本分ではないか。プウィアンヴェナトルは小型の獲物を捕らえる腕前には自信があった。なんとか肉食のポリシーを貫きながら、カルカロ族とは違った方向性の戦略はないものか。
カルカロ族の圧迫を受けるこの地域から、移住も考えていた。はるか北の中国北部には、豊かな森があると聞く。そこでは小型の獲物がふんだんに得られるが、ドロマエオ族、コンプソ族、ティラノ族など競合するハンターも多く、しのぎを削っている。すでに競争が激しい地域に参入するのは難しい。一方、ここから南に向かうと半島や多数の島々がある。そのさらに南には、大きな陸地があるという。そこにはたくさんの竜脚類が繁栄しており、ハンターはほとんどいないと聞いていた。獲物を探しながら、少し南方に移動してみようかな。
(注)プウィアンヴェナトルの気持ちを考えたフィクションです。
コメント ( 0 ) | Trackback ( 0 )
ネオヴェナトルC

これは、お正月にずっとネオヴェナトルBを描いていたら、頭の中にネオヴェナトルが住みついて「縞模様がマンネリだ。もっとかっこいい模様にしろ。」というので。
コメント ( 0 ) | Trackback ( 0 )
ネオヴェナトルB
絵としての難しさ
恐竜の姿形や特徴についての知識とは別に、絵を描く上での技術が要る。
古生物の場合、見たものを正確に描写するのではなく、見たことのないものを想像で描くわけだから、より困難があるはずである。
一頭の恐竜の体にもパースの問題がある。距離と視点を決めて矛盾がないようにする必要がある。
明暗の階調も難しい。光源の位置や散乱の度合いを決める必要がある。見たことのない立体物に陰影をなんとなく描いても、正しいという保証がない。
さらに哲学的で厄介なのは、決まるポーズと決まらないポーズがあることである。動物園で動物が歩き回る様子を、連続写真なり動画のコマ送りで見ると、決まるポーズと決まらないポーズがある。恐竜の復元骨格を色々な角度から撮っても、良いアングルというものが決まってくる。なぜ、決まる角度・ポーズというものがあるのだろうか。図鑑や映画などどこかで見たような、典型的なポーズが「決まるポーズ」と認識されるのだろうか?なぜその構図、アングルが「良い」のだろうか。
そんなことを考えてながら描いている。
コメント ( 0 ) | Trackback ( 0 )
ネオヴェナトルの神経血管網:敏感な吻はスピノサウルスに限らない
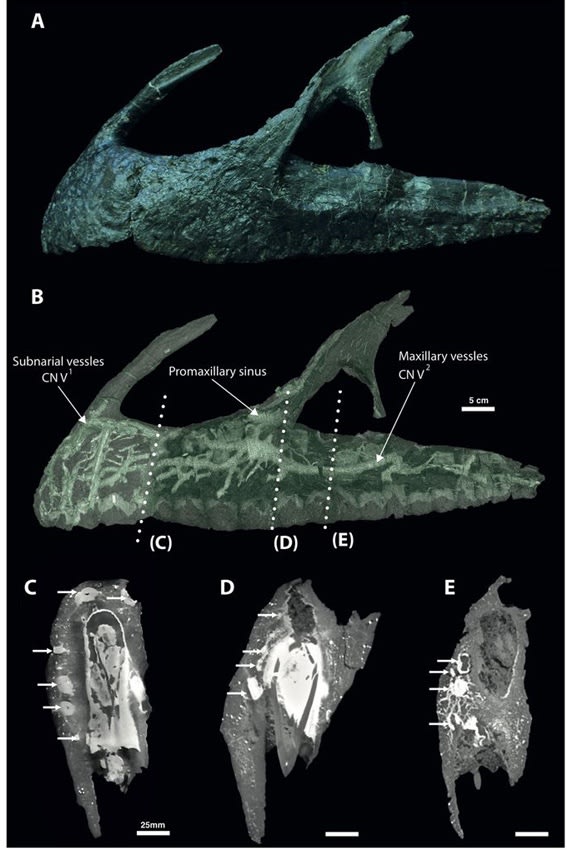
Copyright 2017 Barker et al.
スピノサウルスの半水生適応の研究では、吻部の感覚器の話があった。スピノサウルスの吻(前上顎骨)の表面には多数の孔(神経血管孔)があいており、CTスキャンで内部を観察すると、複雑に枝分かれした神経血管網と思われる管状構造がよく発達している。これは現生のワニと同様に機械受容器と連絡していると考えられ、水中で獲物による振動を感知するなど、半水生適応と関連づけられていた。つまりスピノサウルスのよく発達した神経血管網は、水中で獲物を探すための特殊化と考えられた。バリオニクスについてもそのようにいわれていた。
しかし、本当にスピノサウルス類で発達した形質なのかどうかは、他の獣脚類と比較しないとわからない。実は、スピノサウルス類以外の吻部の神経血管網については、詳細に研究されていなかった。Barker et al. (2017) は、ネオヴェナトルの前上顎骨と上顎骨の表面に、多数の神経血管孔があることに注目して、高精度のCTスキャンで内部構造を研究した。
その結果、ネオヴェナトルの前上顎骨と上顎骨の内部には、スピノサウルスに匹敵するほどよく発達した神経血管網のネットワークが観察された。特に前上顎骨と上顎骨の前半部分で最も発達していて、複雑に枝分かれ、吻合した管状構造がみられた。分岐した枝は最終的に骨表面の神経血管孔に開口していた。
著者らはワニの神経との比較などから、これらの神経血管網を(一部は血管も含まれているが、)主に三叉神経の枝と同定した。前上顎骨には、三叉神経の眼神経CN V1(鼻骨枝の前上顎枝)が分布している。上顎骨には、三叉神経の上顎神経CN V2が分布していて、上顎と歯からの感覚を伝達している。
このようなデータは、吻部の神経の発達に関して、スピノサウルスが獣脚類の中で特別ではないことを示している。典型的な陸上の捕食者であるネオヴェナトルが、スピノサウルスに匹敵する神経血管網と吻部の鋭敏な感覚をもっていることから、これが半水生適応とは考えにくい。ネオヴェナトルが時々、水中の魚などを捕食したことは考えられるが、体の解剖学的特徴などから半水生は支持されない。ネオヴェナトルは他のカルカロドントサウルス類やアロサウロイドと同様に、中型の植物食恐竜を捕食したと考えられている。
スピノサウルスの神経血管網は確かに水中で獲物を検知するのに役立ったかもしれないが、それが水生適応のためだけに発達したというのは言い過ぎであることになる。むしろ、大型の肉食恐竜は一般に、吻部に従来考えられていたよりもよく発達した神経血管網をもっており、獲物の扱いprey handling など他の目的に利用していた可能性が高い。Barker et al. (2017)は3つの目的をあげている。
1つは、肉食恐竜の吻部の鋭敏な感覚は、獲物の解体・処理に用いられたというものである。例えば、骨を避けて肉だけをそぎ取ることが必要になる。ネオヴェナトルの上顎骨歯の表面の微小な摩耗痕が解析されている。そのパターンは、チーターのような骨を噛むことを避ける肉食動物と同じ特徴をもつことから、ネオヴェナトルは骨を避けて肉だけを食べたと推測される。その際に吻部や歯の鋭敏な感覚は、注意深く骨と肉を識別するのに役立ったと考えられる。
2つめは種内のコミュニケーションで、ティラノサウルスなどの大型肉食恐竜は、闘争の際に顔や顎を噛みあうことが知られている。儀式的な闘争などで吻部・顔面の感覚は重要だった可能性がある。
3つめは、巣作り行動の際に穴を掘ったり、適切な温度の場所を探したり、メンテナンスする上でも吻部の鋭敏な感覚は役立っただろうとしている。
確かに吻部の感覚器は、ワニのように水中の感覚に用いるとは限らない。現生鳥類ではキーウィ科、シギ科、トキ科のようにくちばしで餌を探す鳥の吻部に、グランドリー小体やヘルプスト小体という感覚受容器があって、餌を探す際に獲物を認識している。一方オウム類では果実を割るなどの操作に重要であるという。
参考文献
Chris Tijani Barker, Darren Naish, Elis Newham, Orestis L. Katsamenis & Gareth Dyke (2017) Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii. Scientific Reports 7: 3749 DOI:10.1038/s41598-017-03671-3
コメント ( 0 ) | Trackback ( 0 )
アウストラロヴェナトル
大きい画像
ネオヴェナトル科のメガラプトル類アウストラロヴェナトルは、2006年から2010年にかけてオーストラリア恐竜の時代博物館とクイーンズランド博物館の発掘調査により発見され、2009年に記載された。アウストラロヴェナトルの化石は、ティタノサウルス類ディアマンティナサウルスの骨とともに散在して発見された。2009年の論文では、分離した歯、左の歯骨、数本の肋骨、腹肋骨、前肢と後肢の部分骨格が見つかっていた。
その後、前肢や後肢の追加の骨が発見されて、2012年に続報の論文が出ている。新しく見つかった前肢の骨は、左右の上腕骨、右の撓骨、右の撓側骨(手根骨)、右の第1遠位手根骨、右の第I中手骨、左の指骨II-1, II-2, III-3、ほとんど完全な右の指骨II-3(末節骨)である。右の指骨I-2 とIII-4は既に見つかっていたが、今回の追加で第I、第II、第III指の末節骨がそろったことになる。また前肢全体としてもほとんどの骨が見つかっているので、コンピュータ上で復元した像(手)が示されている。
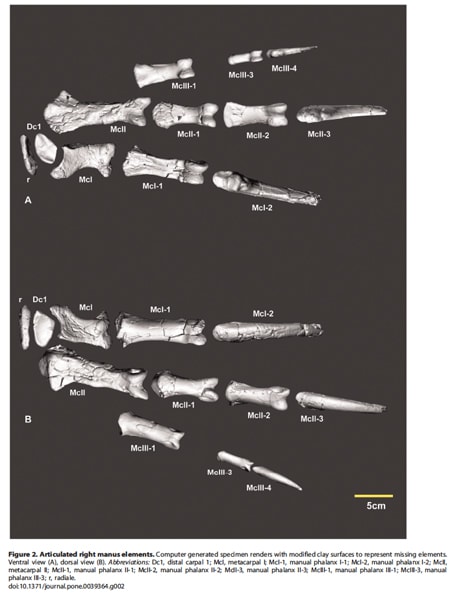
図1 アウストラロヴェナトルの右手の復元(発見されている骨を示す)。Copyright 2012 White et al.
論文の主体をなすresultsの部分は、前肢の骨の形態学的記載が延々と続く。これは研究には絶対不可欠で本質的なことと思うが、アマチュア恐竜ファンにとって面白い部分ではない。興味があるのは、比較して考察している部分である。追加の発見により、アウストラロヴェナトルは四肢の骨についてはネオヴェナトル科の中でも最も完全なものとなったので、他のネオヴェナトル類と比較している。
上腕骨が比較できるのはフクイラプトルとチランタイサウルスしかない。アウストラロヴェナトルの上腕骨は、フクイラプトルと似ている。両者とも骨幹が同じように曲がっており、三角胸筋稜deltopectoral crestは丸く、近位端も遠位端も細くなっている。アウストラロヴェナトルとフクイラプトルでは、上腕骨遠位端のradial condyle とulna condyleの形状が異なるようである。チランタイサウルスの上腕骨はもっとがっしりしており、よりまっすぐである。チランタイサウルスの三角胸筋稜は骨幹から90°近い角度で立ち上がっている。
指骨II-1の形態は、アウストラロヴェナトルとフクイラプトルで異なっている。遠位の関節顆distal condyles と近位端proximal endの高さが、フクイラプトルではほとんど同じであるが、アウストラロヴェナトルでは遠位の関節顆の方が小さいという。
メガラプトル類は、第I指の末節骨が大きな鋭いカギ爪として発達している。メガラプトル、アウストラロヴェナトル、フクイラプトルに共通して、内側と外側で溝の位置が非対称という特徴がある。内側溝medial grooveは先端に向かって中央を通っているが、外側溝lateral grooveは先端に向かって背側を通っている。一方、第II指の末節骨では内側と外側で同様であり、対称的な形をしている。
また、アウストラロヴェナトルでは第I指の末節骨は第II指の末節骨よりもひとまわり大きいくらいで、第I指と第II指でそれほど大きさの差はない。これはフクイラプトルでも同様であるという。一方、メガラプトルでは第I指の末節骨は非常に大きく、第II指の末節骨はそれよりもかなり小さい。つまり第I指と第II指で大きさの差が著しい。
末節骨の基部の高さと幅の比率が、ネオヴェナトル科の共有派生形質として用いられている。それによると、比率が2.3より大きければネオヴェナトル科に含まれることが支持されるという。アウストラロヴェナトルの場合、第I指、第II指、第III指の比率は、それぞれ2.29, 2.22, 2.41であるという。結構微妙な値であるが、論文では、だからどうだというコメントはしていない。このような新しいデータをもとに、基準なども改訂されていくということかもしれない。
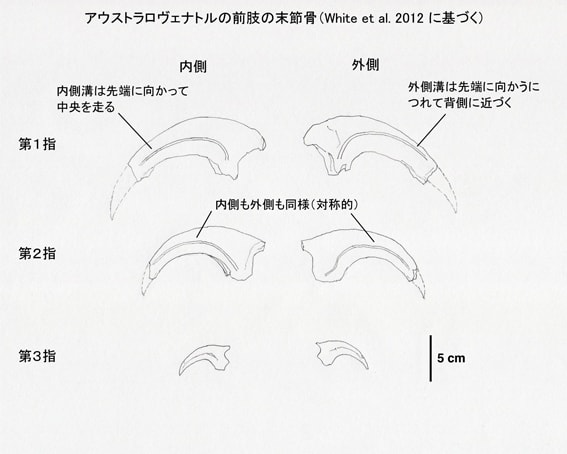
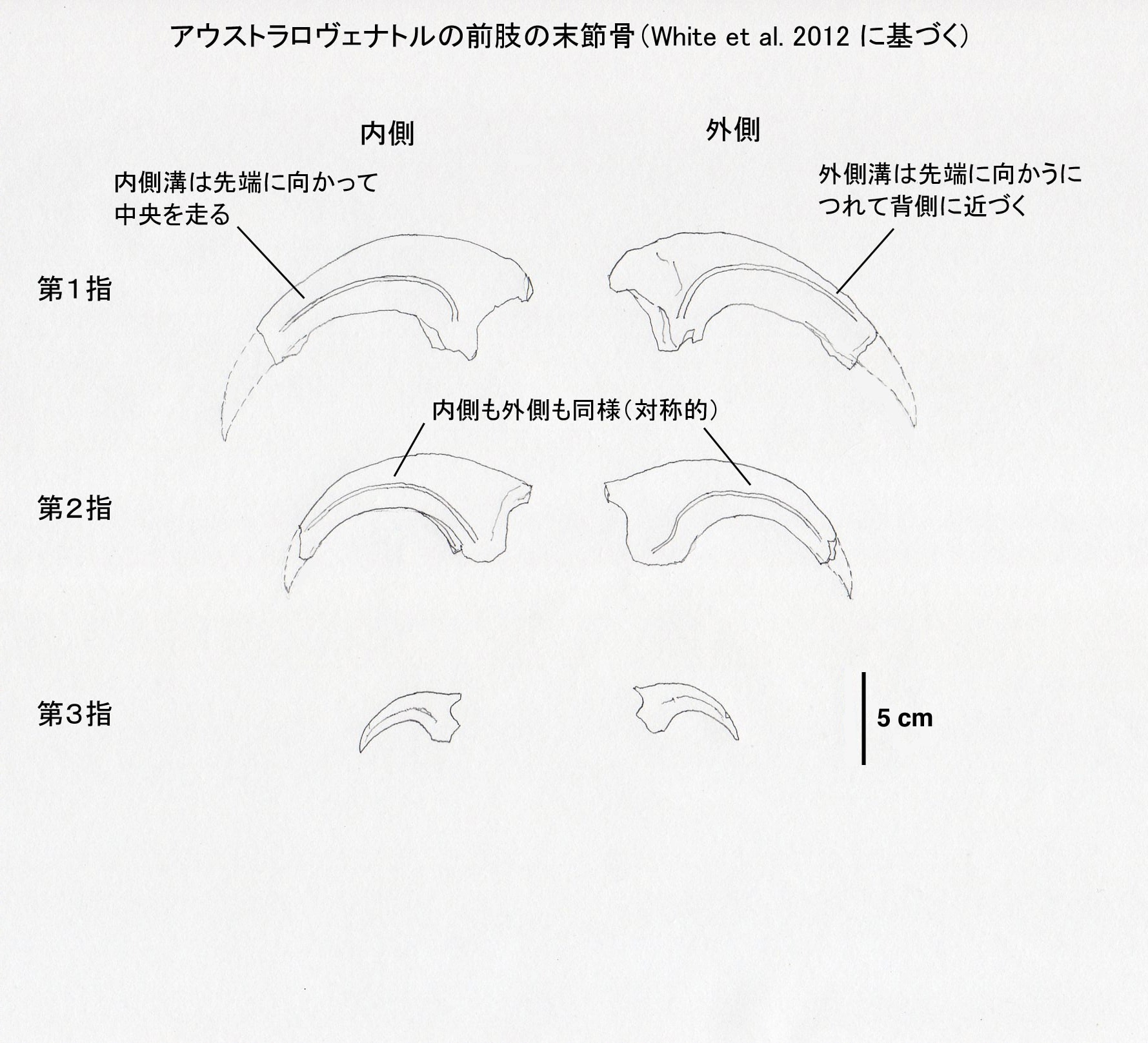
図2 アウストラロヴェナトルの前肢の末節骨。White et al. (2012) の論文の3つの図からスケールを合わせて作成した。カギ爪の先端の推定は筆者による。
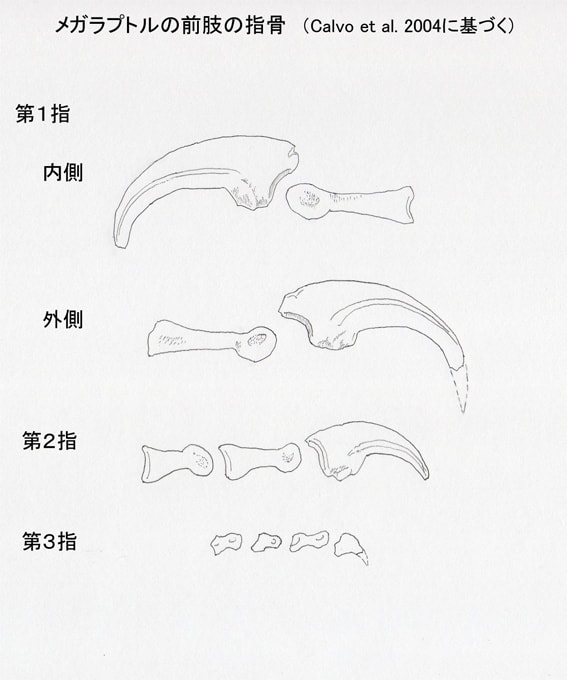
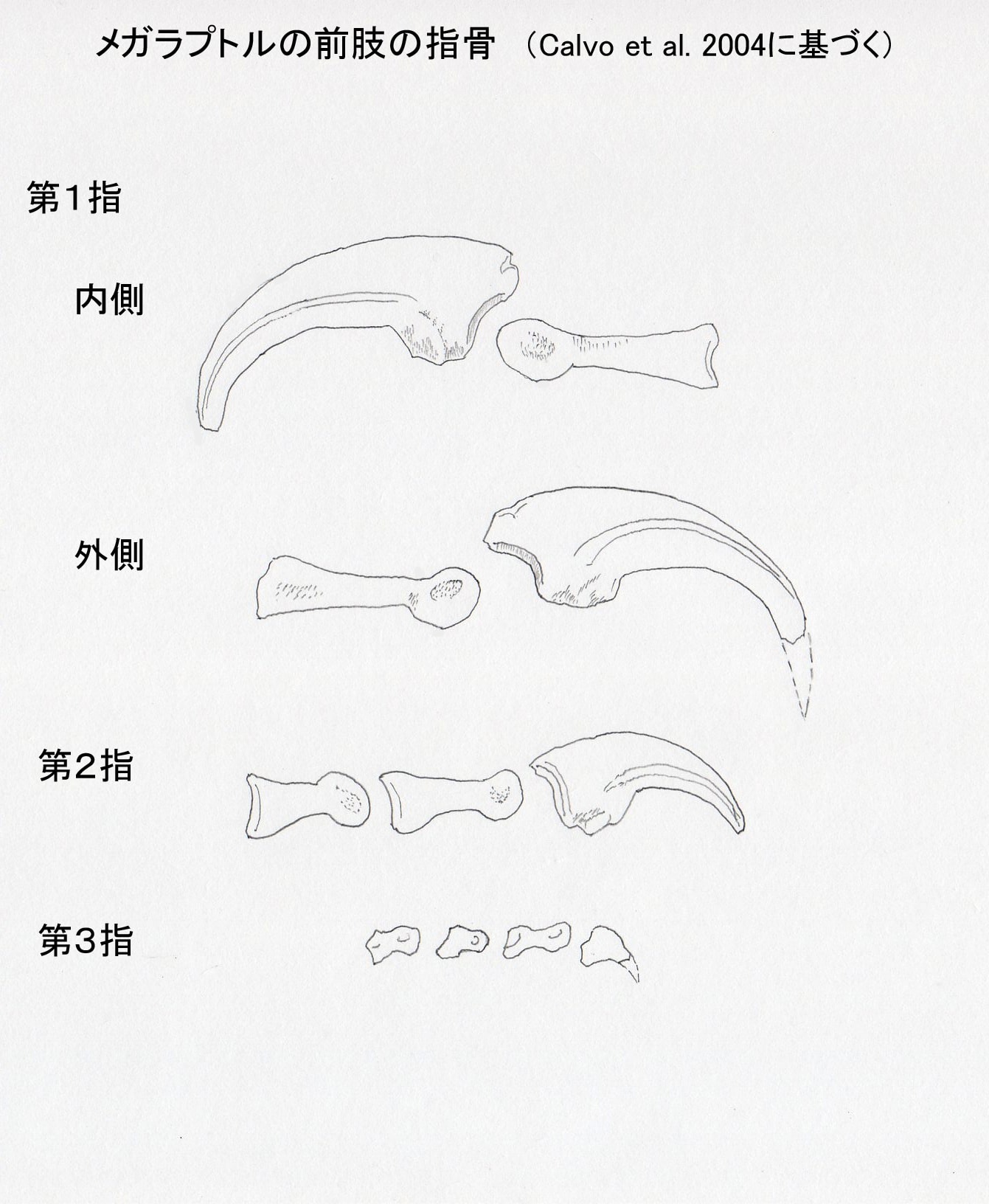
図3 メガラプトルの前肢の指骨。Calvo et al. (2004) に基づいて作成。カギ爪の先端の推定はCalvo et al. (2004)。
残念ながら頭骨はほとんど見つかっていないが、オーストラリアの博物館には復元骨格(全身ではないようだが)があるらしい。これは吻の形や鼻孔が細長い点など、メガラプトルの幼体の復元を参考にしているようにみえる。
参考文献
White MA, Cook AG, Hocknull SA, Sloan T, Sinapius GHK, et al. (2012) New Forearm Elements Discovered of Holotype Specimen Australovenator wintonensis from Winton, Queensland, Australia. PLoS ONE 7(6): e39364. doi:10.1371/journal.pone.0039364
Calvo JO, Porfiri JD, Veralli C, Novas F, Poblete F (2004) Phylogenetic status of Megaraptor namunhuaiquii Novas based on a new specimen from Neuquen, Patagonia, Argentina. Ameghiniana 41: 565–575.
Hocknull SA, White MA, Tischler TR, Cook AG, Calleja ND, et al. (2009) New Mid-Cretaceous (Latest Albian) Dinosaurs from Winton, Queensland, Australia. PLoS ONE 4(7): e6190. doi:10.1371/journal.pone.0006190
コメント ( 0 ) | Trackback ( 0 )
メガラプトル幼体


頭骨の一部は見つかったもののまだまだ不完全であるし、胸郭が大きいなどといわれても全身のバランスがわからないので、復元は依然として難しいと思われる。一見巨大なヴェロキラ風で、よくみると前肢が頑丈でパワフル、後肢と尾はドロマエオでない、といった感じか。最初はカギ爪が後肢のものと思われて巨大なドロマエオサウルス類とされ、その後前肢のカギ爪とわかってメガロサウルス類、カルカロ風、アロ風などに描かれたわけであるが、巡り巡ってユタラプトルなど大型のドロマエオサウルス類に似たイメージに戻ったように思われる。また、近縁のメガラプトル類が皆同じような細長い顔と決まったわけでもないだろう。
Benson et al. のネオヴェナトル科の樹立のときは、ネオヴェナトルとアエロステオンの骨格に類似点が多いことを強調していた(ネオヴェナトルの記事参照)。しかしPorfiri et al. (2014) の説ではネオヴェナトルだけがアロサウルス上科に取り残され、メガラプトル類はごっそりティラノサウルス上科にいっているわけである。この辺りはどうなのだろうか。また、Porfiri et al. (2014)はチランタイサウルスとシアッツはメガラプトル類ではないと考えている。一方オーストラリアの研究者はネオヴェナトル科を支持しているようである。
しかし南米産はやはり侮れませんね。他にもウネンラギア類などの曲者もいるし。
コメント ( 0 ) | Trackback ( 0 )
メガラプトルの頭骨

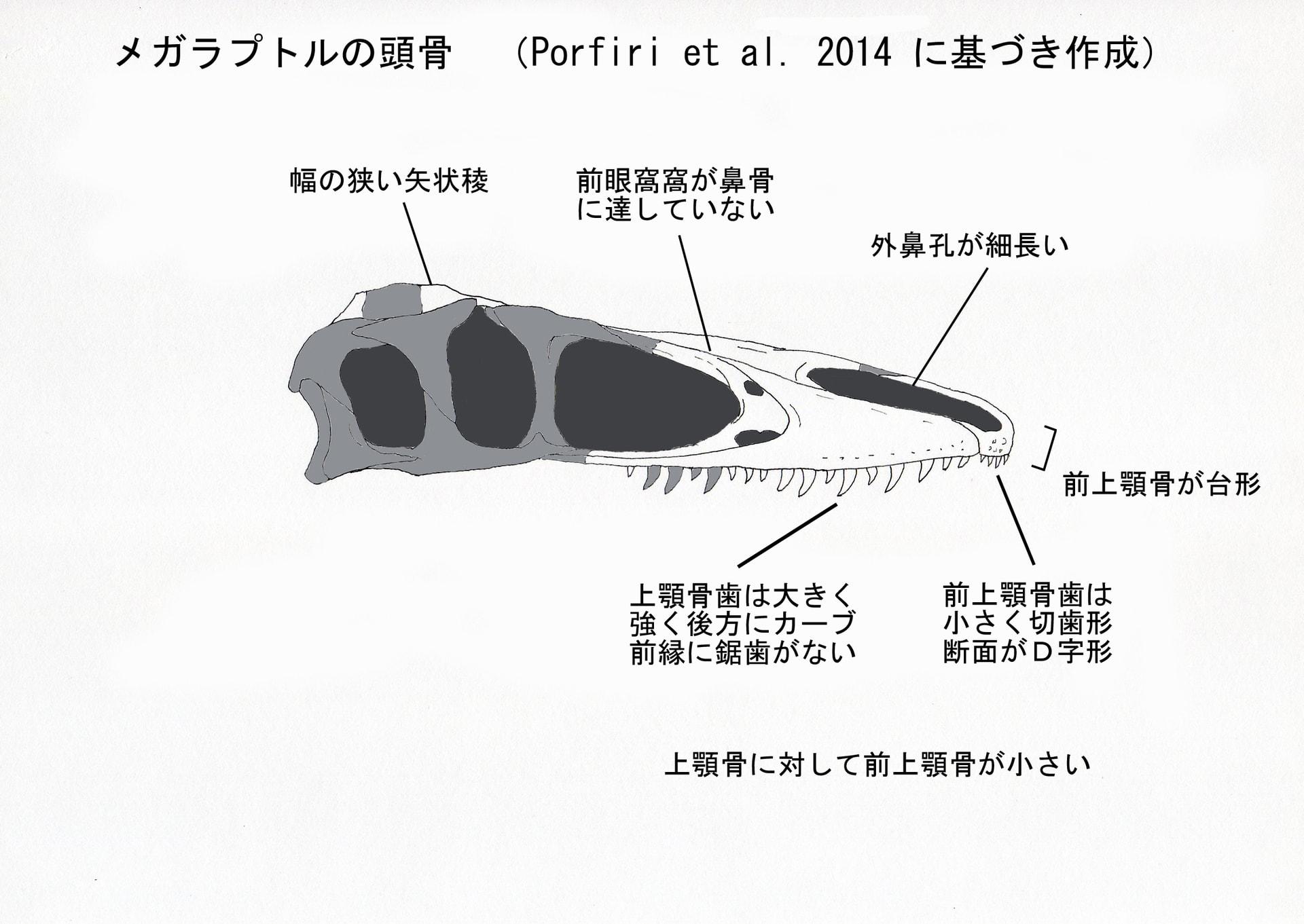
大きい画像
メガラプトル類は、前肢の第1指と第2指に大きく発達した、幅が狭く鋭いカギ爪をもつことを特徴とする獣脚類である。派生的なメガラプトル類(メガラプトル科Megaraptoridae, Novas et al. 2013)は白亜紀のセノマニアンからサントニアンのゴンドワナ地域から知られている。メガラプトル類の系統的位置については近年、議論が続いており、研究者によって基盤的なコエルロサウルス類、基盤的なテタヌラ類(メガロサウロイド)、カルカロドントサウルス類に近縁なアロサウロイドなどに分類されてきた。最近、Novas et al. (2013)はコエルロサウルス類、なかでもティラノサウルス上科に含まれるとした。
このように解釈が分かれる理由はメガラプトル類の骨格が不完全なためで、特に頭骨の骨はほとんど発見されていなかったため、頭骨の形態についてはきわめて情報が乏しかった。Porfiri et al. (2014) は、部分的な頭骨と胴体の骨格を含む、メガラプトルの幼体の標本について記載している。この標本によって初めて、メガラプトルの頭骨の形態についてかなりの情報が得られたわけである。
この標本MUCPv 595はアルゼンチンのネウケン州Baal quarryで、白亜紀後期チューロニアン-コニアシアンのポルテスエロ層Portezuelo formation から発見された。この標本は、左右の前上顎骨、上顎骨、鼻骨、左の前頭骨、部分的な脳函、関節した9個の頸椎、胴椎1-8, 10-12, 仙椎3-5, 4個の近位の尾椎、腹肋骨、肩甲骨、烏口骨、上腕骨、部分的な恥骨からなる。
この標本は肩甲骨、恥骨、頸椎などの形態が、これまで見つかっているメガラプトルの化石と一致した。ただし大きさは小さく、メガラプトルの成体では推定全長9-10 m とされるのに対してMUCPv 595は推定全長3 m と考えられた。また、神経弓と椎体など、いくつかの骨が癒合していないことからも幼体と考えられる。さらに肩甲骨と恥骨について組織切片を作成し、成長線の数が少ないことも確認している。
この標本は幼体なので、頭骨のプロポーションや隆起の発達程度が成体と異なるかもしれない。しかしそれらを考慮しても、アロサウロイドのような基盤的なテタヌラ類とは多くの違いがあり、一方で基盤的なティラノサウロイドと多くの類似点があるといっている。
前上顎骨の外表面にはいくつかの大きな孔foramenがあいているが、これは多くのティラノサウロイドにみられる。前鼻孔突起prenarial processは棒状で非常に長く伸びており、前上顎骨体の長さの300%に達する。そのため、外鼻孔がきわめて前後に長い。これはキレスクス、プロケラトサウルス、ディロング、オルニトレステス、グァンロンと同様である。
前上顎骨の前縁は前背方に傾き、後縁は後背方に傾いているので、前上顎骨は側面からみて下が短い台形の輪郭trapezoidal contourをなす。つまり前上顎骨の腹側がより強く短縮している。この台形の輪郭は、プロケラトサウルス、グァンロン、ディロング、いくつかの派生的なティラノサウルス類(ティラノサウルス、タルボサウルス)と似ている。ネオヴェナトルやアクロカントサウルスのようなアロサウロイドにはこの台形の輪郭はみられない。
メガラプトルの前上顎骨体は、上顎骨の長さに対して相対的に小さい。この点はアロサウロイドとは顕著に異なっている。プロケラトサウルス、ディロング、キレスクス、派生的なティラノサウルス類(ティラノサウルス、タルボサウルス)のようなティラノサウロイドは皆、非常に短い前上顎骨をもっている(グァンロンはやや長いらしい)。(注:論文ではメガラプトルの前上顎骨体は上顎骨の長さの0.7%となっているが、どうも数値が疑問である。7%の誤りか。)
上顎骨は側面からみて三角形に近く、前方突起rostral ramusは強く前後にのびて、背側縁はほとんど直線状になっている。この形状は基盤的なテタヌラ類とも、基盤的なコエルロサウルス類とも、基盤的なティラノサウロイドとも異なっている。この点では、メガラプトルはオルニトミムス類、トロオドン類、派生的なティラノサウルス類と似ている。
前眼窩窩はよく発達している。前眼窩窩には2つの孔があり、前後方向にはほぼ同じ位置で、上下に並んでいる。背側の孔はmaxillary fenestra で、腹側の孔はpromaxillary foramenかもしれないという。しかしこのような孔の配置は、他の獣脚類にはみられない独特のものである。
口蓋骨突起palatal processは細長く、アリオラムスやキレスクスと似ているが、アロサウルス、シンラプトル、マプサウルスなどにみられる太い状態とは異なっている。シンラプトルより派生的なアロサウロイドでは歯間板interdental plateが癒合しているが、メガラプトルでは歯間板が癒合していない。
左右の鼻骨は癒合していない。メガラプトルの鼻骨は細長くのびており、その横幅は全長にわたって大体一様である。これは、アクロカントサウルス、カルカロドントサウルス、オルニトミムス類、トロオドン類、ドロマエオサウルス類と似ている。メガラプトルの鼻骨は、ティラノサウルス類の鼻骨とは特に似ていない。基盤的なアロサウロイドでは鼻骨の前端(外鼻孔の周り)にはっきりしたnarial fossa があるが、メガラプトルにはnarial fossaはない。また、アロサウロイドや他の基盤的テタヌラ類に特徴的な、鼻骨の側方縁の粗面rugosityはみられない。アロサウロイドなどの基盤的テタヌラ類と異なり、メガラプトルでは前眼窩窩が鼻骨に達していない。
前頭骨は四角形で、上側頭窩supratemporal fossa が広く拡張している。そのため前頭骨の後半で、幅の狭い矢状稜sagittal crest が形成されている。このことから、保存されていない頭頂骨にも幅の狭い矢状稜があったことが示唆される。このような特徴は、ティラノサウロイドの状態に似ている。アロサウロイドでは、上側頭窩の拡張は中くらいで、矢状稜はより幅広い。またメガラプトルの前頭骨は前半が低く後半が高くなっていて、中ほどに段差がある。
脳函についても詳細に観察しており、いくつかの形質はアロサウロイドよりもコエルロサウルス類と似ているといっている。大後頭孔は比較的大きく、後頭顆の頸部の基部に深く達している。一方、基盤的な獣脚類では大後頭孔は比較的小さく、後頭顆の頸部の基部にわずかに達しているか(アクロカントサウルス、シンラプトル)、または達していない(メガロサウルス類)。傍後頭骨突起は多くの獣脚類と同様に後側方かつ水平に突き出しており、アロサウロイドに特徴的にみられるような下方に曲がった形ではない。
メガラプトルの上顎には、少なくとも前上顎骨に4本、上顎骨に15本(おそらくは17)の歯があったと考えられる。前上顎骨歯は切歯形incisiviformで小さく(歯冠長10 mm)、大きな上顎骨歯(17 mm)と対照的である。前上顎骨歯は、ティラノサウルス類にみられるようなD字形の断面をしている。上顎骨歯は強く後方にカーブしていて、前縁には鋸歯がなく、両側から凹んだ8字形の断面をなす。
Porfiri et al. (2014) は、Novas et al. (2013)のデータに新しいメガラプトルの標本から得られた情報を加えて、再び系統解析を行った。その結果やはり、メガラプトル類Megaraptoraはコエルロサウルス類の中でティラノサウルス上科に含まれることが支持された。ここではメガラプトル類がティラノサウロイドと共有する派生形質は、1)前上顎骨歯が切歯形incisiviformで、2)断面がD字形、3)前上顎骨の外側面にいくつかの大きな円形の孔がある、4)外鼻孔の長さが幅の3倍以上ある、5)歯骨の最初の歯槽が後方の歯槽よりもかなり小さい、6)頭頂骨に正中の矢状稜sagittal crestがある、つまり左右の上側頭窩の間の部分の幅が非常に狭い、などである。
この分岐図ではMegaraptoraのうちフクイラプトルが最も基盤的なもので、それ以外の派生的なメガラプトル類、メガラプトル科Megaraptoridaeにはアウストラロヴェナトル、アエロステオン、オルコラプトル、メガラプトル、そして意外なことにエオティランヌスが含まれた。
著者らはこれまで得られたメガラプトル科の標本から、派生的なメガラプトル類の骨格を復元することができるという。これらの動物は、細長い吻と小さめの歯のある長い頭骨をもっていた。首もまた華奢で、よく発達したS字形の頸椎をしていた。肋骨と腹肋骨の形から、胸郭は非常に幅広く丈も高かった。胸帯は頑丈で、長く太い前肢を支え、手の第1指と第2指には大きく鋭いカギ爪があった。後肢は華奢でスレンダーだった。このような体形は他の獣脚類にはみられないユニークなものであるという。
Novasらはさらに、最近のチレサウルスの論文の中でも広汎な系統解析を行っている。その結果今度は、メガラプトルはティラノサウルス上科の中ではなく、最も基盤的なコエルロサウルス類と位置づけられた。どうも未だにpendingという状況のようである。
確かに、ネオヴェナトルとはかなり異なる顔つきのようである。頭骨復元図は吻が細長く、トロオドンやドロマエオ系、あるいはアリオラムス的な顔にみえる。ティラノサウロイドかどうかわからないが、コエルロサウルス類の一種のような雰囲気はある気がする。そうすると近縁のアエロステオンも、従来は羽毛の生えたアロサウルスそのものの姿に描かれてきたが、今後は復元像が変わってくるのかもしれない。さらにはフクイラプトルのイメージにも影響してくるだろう。
参考文献
Porfiri, J.D., Novas, F.E., Calvo, J.O., Agnolín, F.L., Ezcurra, M.D. & Cerda, I.A. (2014) Juvenile specimen of Megaraptor (Dinosauria, Theropoda) sheds light about tyrannosauroid radiation. Cretaceous Research 51, 35-55.
Novas, F.E., Agnolín, F.L., Ezcurra, M.D., Porfiri, J. & Canale, J.I. (2013) Evolution of the carnivorous dinosaurs during the Cretaceous: the evidence from Patagonia. Cretaceous Research 45, 174–215.
Fernando E. Novas, Leonardo Salgado, Manuel Suarez, Federico L. Agnolin, Martin D. Ezcurra, Nicolas R. Chimento, Rita de la Cruz, Marcelo P. Isasi, Alexander O. Vargas & David Rubilar-Rogers (2015) An enigmatic plant-eating theropod from the Late Jurassic period of Chile. doi:10.1038/nature14307
コメント ( 2 ) | Trackback ( 0 )
オーストラリアのネオヴェナトル類
英国のBensonとオーストラリアのRich, Vickers-Richらの共同研究で、オーストラリア南部の獣脚類化石についてレビューした論文が出ている。オーストラリア南部ヴィクトリア州のOtwayおよび Strzelecki層群(白亜紀前期アプト期からオーブ期)からは、少なくとも37の分離した骨と、多数の歯が発見されている。その中にはアロサウロイド、ティラノサウロイド、スピノサウルス類の亜成体、ケラトサウルス類、オルニトミモサウルス類、基盤的コエルロサウルス類、3種類のマニラプトル類が含まれ、全部で9種類もの獣脚類がいたと考えられるという。
論文の主旨としては、およそ以下のようである。オーストラリアはゴンドワナ地域なので、これまで白亜紀の恐竜相は南アメリカや南極などと関連があると考えられてきたが、最近の研究ではむしろローラシアの恐竜相と似ていることがわかってきた。今回の獣脚類の構成をみると、アロサウロイドと基盤的コエルロサウルス類(ティラノサウロイドやオルニトミモサウルス類を含む)が多いこと、アベリサウルス類のようなケラトサウルス類が少ないことなど、同時代のアフリカや南アメリカ(高緯度で乾燥した環境)とは大きく異なっており、むしろ温帯のローラシア地域と共通点が多い。多くの恐竜のグループ(高次分類群)は、パンゲアの分裂以前のジュラ紀には汎世界的に分布しており、従来「ゴンドワナ的」とされた恐竜相の特徴の一部は、大陸の分裂によりもたらされたものではなく、気候の違いによる地域性を反映しているのかもしれない、と考察している。
ここでは論文で記載された中から、ネオヴェナトル類の化石だけをピックアップして、このような断片的な骨がどうしてネオヴェナトル類と同定されたのか、参考のためにメモしておきたい。
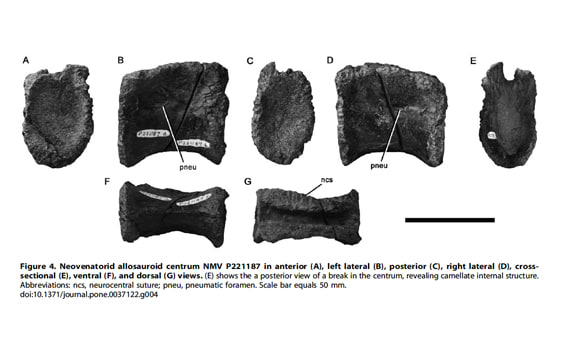
Copyright: 2012 Benson et al.
NMV P221187は、胴椎の椎体で、長さが高さよりも少し大きい(長さ60mm, 高さ48mm)。椎体の関節面は高さが幅の約1.25倍で、弱く凹んでいる。神経弓との結合面の形状から、神経弓と椎体は癒合しておらず、幼体と思われる。椎体の外側面は腹側で合して、低く角張った縦方向の腹側稜ventral ridgeをなしている。含気孔は椎体側面の前背方にあり、前後に長い楕円形をしている。また含気孔の腹側縁は背側縁よりも突出しているので、孔は側方というよりも背側方を向いて開いている。
椎体を切断して断面をみると、腹側や背側に多数の小室camellaeがある。このようなcamellate内部構造は、ケラトサウルス類、カルカロドントサウリア、ティラノサウルス科、オヴィラプトロサウルス類、テリジノサウルス類にみられる。しかしケラトサウルス類では椎体の側面に2個の含気孔があるが、この椎体では1個である。前後に長い、背側方に開いた含気孔が椎体側面の前方にあるという点で、この椎体はカルカロドントサウリアとティラノサウルス科のものに最もよく似ている。
また、低く角張った縦の腹側稜は、ネオヴェナトル類であるネオヴェナトルの第6、7胴椎とアエロステオンの第6~8胴椎にもみられる。このことと含気孔の形態、camellate内部構造から、NMV P221187 はNeovenatoridae indet.とされた。
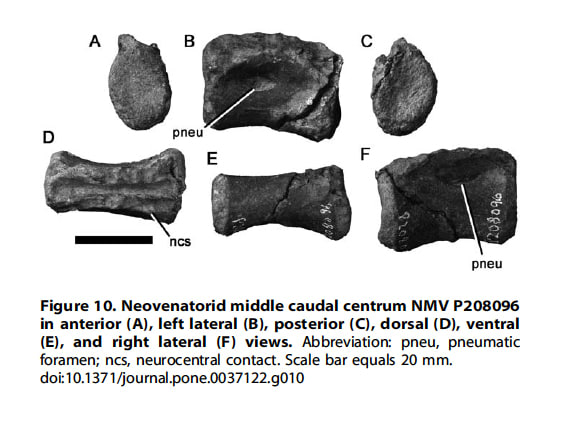
Copyright: 2012 Benson et al.
NMV P208096は椎体で、比較的前後に長い形状から、中央部の尾椎と同定された。神経弓と椎体は癒合しておらず幼体と考えられる。横方向につぶれているので正確にはわからないが、関節面は高さが幅より少し大きいと思われる。椎体の腹側面は一様に丸く、稜や溝などはない。大きく前後に長い楕円形の含気孔が、椎体側面の中央より前方に位置している。
獣脚類の中央の尾椎にはもともと含気孔はなく、メガラプトル類、オヴィラプトロサウルス類、テリジノサウルス類で独立に獲得されている。しかしオヴィラプトロサウルス類の尾椎は短く、この尾椎とはプロポーションが異なる。 テリジノサウルス類では含気孔が小さく、最も近位の尾椎以外では神経弓にある。これらの違いと、メガラプトル類アエロステオンの中央の尾椎と似ていることから、NMV P208096 はNeovenatoridae indet.とされた。
Copyright: 2012 Benson et al.
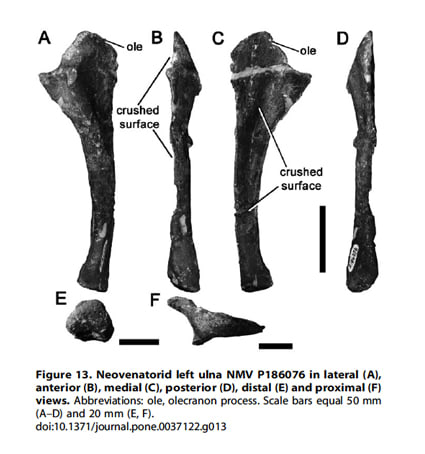
NMV P186076は左の尺骨で、Rich & Vickers-Rich (2003)により報告され、Smith et al. (2008) によってcf. Megaraptorとして記載された。最近のほとんどの研究は、NMV P186076はメガラプトルに近縁のアロサウロイドとしている。NMV P186076は、アウストラロヴェナトルやメガラプトルのような他のメガラプトル類の尺骨と多くの類似点を示している。基盤的なネオヴェナトル類の尺骨は知られていないので、いまのところNMV P186076はメガラプトル類と特定することはできず、ここではNeovenatoridae indet.とされる、という。
Copyright: 2012 Benson et al.
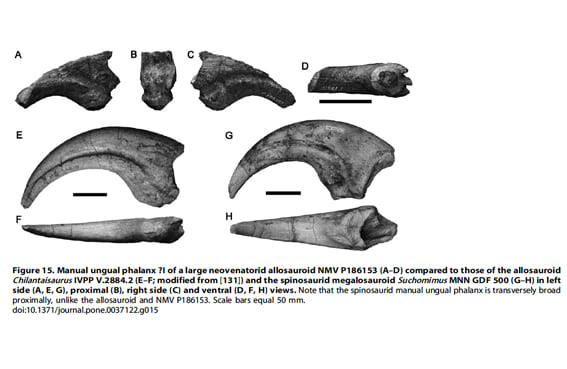
NMV 186153は、大きな前肢の末節骨で、遠位端と近位背側が欠けている。保存された部分の大きさはバリオニクスやアロサウルスの第1指の末節骨に匹敵するもので、全長8~9mの大型獣脚類であることが示唆される。NMV 186153の近位端は、保存された最も遠位の部分(折れる前の末節骨の中央あたりに相当)と比べてわずかしか幅広くない。これは比較的幅の狭い(うすい)爪をもつチランタイサウルスのようなネオヴェナトル類の状態に似ている。また、近位部の幅が広く遠位に向かってかなり細まっている、アロサウルスのようなより基盤的なアロサウロイドやスピノサウルス類を含むメガロサウロイドのものとは異なっている。しかし、NMV 186153はメガラプトル類の末節骨ほど幅が狭くはないようであり、ここではネオヴェナトル類とされる(メガラプトル類ではないかもしれない)。
オーストラリアのネオヴェナトル類としては既にアウストラロヴェナトルの部分骨格が知られているが、当然他の種類もいただろう。これらの断片的な骨も別種であってほしいものである。
参考文献
Benson RBJ, Rich TH, Vickers-Rich P, Hall M (2012) Theropod Fauna from Southern Australia Indicates High Polar Diversity and Climate-Driven Dinosaur Provinciality. PLoS ONE 7(5): e37122. doi:10.1371/journal.pone.0037122
Copyright: 2012 Benson et al.
コメント ( 4 ) | Trackback ( 0 )
シアッツ・ミーケロルム
CNN日本版はシアッチといい、AFP日本版はシアットと書いている。綴りはシアッツじゃないの?と思ったあなたは正しい。シアッツとは、ユタ州の先住民ユート族の神話に登場する人食い怪物の名であるが、しばしばSee-atchとも綴るとあるから、現地の発音は「シアッチ」に近いのかもしれない。しかし、日本人がこのローマ字綴りを見た時に、普通はシアッチとは読めないと思うので、ここではシアッツとする。
シアッツは、白亜紀後期の初めセノマン期(Cedar Mountain Formation)に、米国ユタ州に生息した大型のアロサウロイドで、北アメリカ初のネオヴェナトル類として2013年に記載された。完模式標本は部分骨格で、いくつかの脊椎骨、腰帯、後肢の骨からなる。ネオヴェナトル類は頭骨が見つからないというお約束でもあるかのように、シアッツも頭骨や前肢などは発見されていない。そのため残念ながら正確な復元は無理である。プレスリリースされた想像図のように、なんとなくアロサウルス風の姿をイメージするしかない。
シアッツに固有の形質は、近位の尾椎に前後に広がったcentrodiapophyseal laminaeをもつが、よく発達したinfradiapophyseal fossaeを欠いていること、前方の胴椎が前後に長いこと、胴椎の神経棘が短く幅広いこと、遠位の尾椎の腹側面が横方向に扁平で前後方向に凹んでおり、断面が亜三角形であること、腸骨の恥骨柄pubic peduncleの寛骨臼縁が横方向に凹んでいること、など非常に難しい。
おそらく5番目の胴椎は、ほとんど完全に保存されている。そこには、ネオヴェナトルやアエロステオンと同様に、低いが明瞭な腹側稜ventral ridgeがあり、関節面は幅よりも高さが大きく、後関節突起にはフランジ状の側方延長部がある。神経弓の基部は丈が高いが、アクロカントサウルスや他のカルカロドントサウルス科の胴椎と異なり、神経棘は短く幅が広い。(想像図を描く上で参考になるのはせいぜいこのくらいか。神経棘が低め、である。)
系統解析の結果、主に胴椎や仙椎の椎体の側腔pleurocoelや、camellate(蜂の巣状)内部構造などの含気性の発達程度や形状から、シアッツは広義のカルカロドントサウルス類(カルカロドントサウリア)とされた。
また1)胴椎の後関節突起のつまみ状tab-likeの延長部や、2)前関節突起の外側に貫通した含気腔、3)尾椎の神経弓の顕著なcentrodiapophyseal laminae、4)腸骨の前寛骨臼突起にcuppedicus shelfがあることなどから、シアッツはネオヴェナトル科Neovenatoridaeに位置づけられた。さらに、「尾椎の神経弓における深いinfradiapophyseal fossaに支えられた顕著なcentrodiapophyseal laminae」により、ネオヴェナトル科の中でもメガラプトル類Megaraptoraに属することが支持されるとある。(シアッツの固有形質には、よく発達したinfradiapophyseal fossaeはないとあるが、メガラプトル類の中ではアエロステオンやメガラプトルに比べてシアッツでは浅いという意味か。二次的に逆転という意味か。)
北アメリカの恐竜相では、白亜紀前期ごろに頂上捕食者top predatorである大型肉食恐竜の構成が大規模に変化したことはよく知られている。ジュラ紀後期にはメガロサウルス類、アロサウルス類、ケラトサウルス類など様々な系統の中型から大型の獣脚類が共存していたが、白亜紀末(カンパニア期以後)になるともっぱら大型のティラノサウルス類で占められるようになる。しかしその間の大型捕食者の化石記録は乏しく、7000万年近いギャップがあるために、この大変化の時期や様相は不明のままであった。例えば白亜紀前期の大型肉食恐竜としては、アクロカントサウルスが知られているくらいであった。
大型のネオヴェナトル類シアッツの発見により、白亜紀中頃の生態系はジュラ紀後期とも白亜紀末とも異なる、ユニークなものであることが示唆された。つまり、白亜紀中頃の頂上捕食者(大型肉食恐竜相)は派生的なカルカロドントサウリアン(カルカロドントサウリアのメンバー)で占められていたと考えられる。よって北アメリカの頂上捕食者は、少なくとも3段階の変遷を経ていたと思われる。
アジアではシャオチーロンの研究により、白亜紀後期に入ってもカルカロドントサウリアンが優勢で、そのためにティラノサウルス類の台頭は遅かったという考えが既に提唱されている。シアッツの発見により、北アメリカでも同様にカルカロドントサウリアンが頂上捕食者の地位を占め続けており、おそらくそのためにティラノサウルス類は大型化できなかったことが推測される。さらに、シアッツの発見された地層からは小型のティラノサウロイドと推定される化石が見つかっており、大型のアロサウロイドと小型のティラノサウロイドが同じ地域に共存していた証拠となることが期待される。
ちなみにネットニュースなどの小見出しで「ティラノと共存?」のようなものが見られたが、よく知らない人には誤解を招くのでよろしくない。「ティラノサウルス」と「ティラノサウルス類」は全く意味が異なるのに、書き手自身が深く理解していないのか、読者の目を引くためなのか。最初から適切な書き方をすべきである。
参考文献
Zanno, L. E. and Makovicky, P. J. (2013) Neovenatorid theropods are apex predators in the Late Cretaceous of North America. Nat. Commun. 4:2827 doi: 10.1038/ncomms3827.
コメント ( 3 ) | Trackback ( 0 )
| « 前ページ |