獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2017年3月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
エウストレプトスポンディルス


大きい画像
舌を噛みそうな名前の言いにくさには定評があるが、長年の恐竜ファンなら記憶にあるはずである。BBCの最初のWalking with Dinosaursで、「海を渡る恐竜」として登場し、リオプレウロドンに海中に引きずり込まれる役も演じている。イギリスの恐竜ファンにはなじみがあるのだろうか。
エウストレプトスポンディルスは、ジュラ紀中期カロヴィアン後期(Oxford Clay Formation)にイギリスのオックスフォードシャーに生息したメガロサウルス類である。非常に古くから知られていた恐竜で、Phillips (1871) によって最初に報告され、その後ストレプトスポンディルス・キュヴィエイと呼ばれたこともあったが、Walker (1964) によってエウストレプトスポンディルス・オクソニエンシスと命名された。しかし何人かの研究者に取り上げられてはきたものの、完全な記載はなされていなかった。Sadleir et al. (2008) によってようやく現代的なモノグラフが出版された。
ホロタイプOUMNHJ.13558 は部分的な頭骨と胴体の骨格で、かなりの部分が見つかっているともいえる。特に脊椎は、頸椎から仙椎までほぼ連続的に保存されている(個々の骨は不完全)。また後肢の骨は比較的保存されている。
ホロタイプは、以下の骨からなる。左右の前上顎骨、上顎骨、前頭骨、歯骨、後眼窩骨、右の頭頂骨、左の鱗状骨、右の涙骨、右の方形骨、部分的な脳函、第2頸椎から第5尾椎までほとんど連続した脊椎と、連続しない後方の尾椎、右の肩甲骨と上腕骨、左右の座骨と恥骨、右の腸骨、左右の大腿骨、脛骨、腓骨、距骨、左右の足の中足骨II, III, IV と部分的な趾骨である。
エウストレプトスポンディルスの固有形質は、涙骨の浅い含気孔lachrymal fenestra の中にもう一つの小さい孔があること、鱗状骨にフランジが発達していて下側頭窓の後背方部の上に覆いかぶさっていること、頸椎から胴椎までの椎体に腹側のキールがないこと、第10頸椎の腹側面の前方部に顕著な凹みがあること、である。さらに他のすべてのスピノサウロイドと区別される特徴として、後眼窩骨の腹側突起の後ろ側に窪みがあり、下側頭窓の前縁を取り囲むような形をなしている。

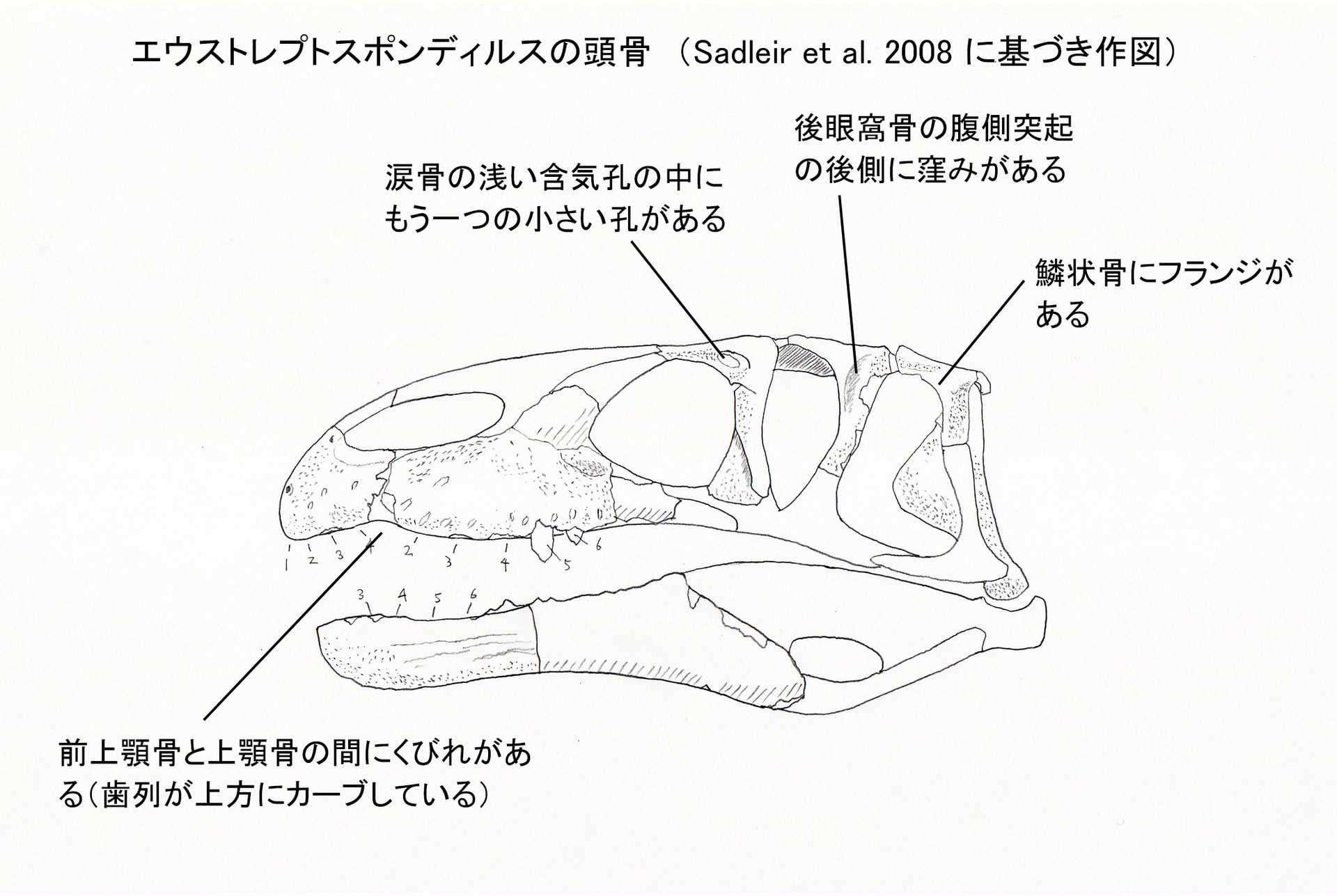
大きい画像
左右の前上顎骨は、どちらも内側の関節面はよく保存されているが、外鼻孔の前後の突起(鼻骨突起と鼻孔下突起)は折れている。右の前上顎骨の3番目の歯槽には完全な置換歯が保存されている。前上顎骨は側面からみると、前後の長さが高さよりも少し大きく、背側縁は凸型にカーブし腹側縁はまっすぐである。背側縁は後背方に向かって、水平から45°の角度をなしている。外側面は前方では凸型で、前方と腹方に神経血管孔が集中している。外鼻孔の周り(下)の部分は凹んでいる。
右の上顎骨は左よりも完全であるが、どちらもあまり保存はよくないようで、前上顎骨との関節面や腹側縁は破損している。右側の方が上方突起と頬骨突起が保存されているが、それらも途中で折れている。右の上顎骨にはひどく破損した5番目と6番目の歯冠がある。その他、左右ともいくつかの歯槽に置換歯が残っている。
上顎骨は前後に長く、前方突起は丈が高く頬骨突起は丈が低い。上方突起は、およそ5番目の歯槽の位置から立ち上がっている。上顎骨には少なくとも9個の歯槽があり、歯槽の形は前後に長い卵形である。3番目の歯槽が最も大きく、それより後方では次第に小さくなる。上顎骨の外側面は平面的で、上方突起の基部に窪んだ前眼窩窩がある。歯列のある腹側縁は前方で少しカーブしており、曲線の頂点は3番目の歯槽の位置にある。
恐竜ファンにとって、外観の「顔つき」に影響するような情報は重要だろう。Walking with DinosaursのCG、及びオックスフォード大学自然史博物館にある頭部模型ではアロサウルスと大差ない印象だったが、Sadleir et al. (2008)の復元頭骨図はもう少しメガロサウルス類らしい、あるいはスピノサウロイドな顔である。特に、吻の前方にはスピノサウルス類やディロフォサウルスのような「くびれ」がある。ただし、関節状態ではなく、前上顎骨も上顎骨も関節面は破損しているとあるので、この組み合わせ方は本当に適切なのか心配になる。
本文を読むと一応根拠があることがわかる。前上顎骨の鼻孔下突起は、上顎骨の前背方面の上にのびているはずであり、そのように前上顎骨と上顎骨を合わせると、上顎骨の歯列の上の窪みと、前上顎骨の4番目の歯槽の上の窪みが一致するという。このように配置すると、前上顎骨と上顎骨の間の部分で、歯列のラインが少しくびれている(凹型にカーブしている)ことになる。他のメガロサウルス類にはこのようなくびれは知られていない。
Sadleir et al. (2008) の系統解析では、エウストレプトスポンディルスはスピノサウルス類と姉妹群をなした(エウストレプトスポンディルス+スピノサウルス類がクレードとなった)。つまり他のメガロサウルス類よりも、スピノサウルス類の祖先と近縁ということになる。その文脈で、スピノサウルス類の進化の過程では、顕著な吻の伸長に先立って、上顎歯列の上方へのカーブ(くびれの位置で)が発達したと考察している。
しかし他の研究、例えばBenson (2010) の系統解析では、エウストレプトスポンディルスはメガロサウルス類の中で基盤的な位置にきている。上顎歯列のカーブはメガロサウルス類とスピノサウルス類の共有派生形質で、その後エウストレプトスポンディルス以外のメガロサウルス類では失われたか、あるいはエウストレプトスポンディルスとスピノサウルス類で独立して獲得されたか、ということになる。

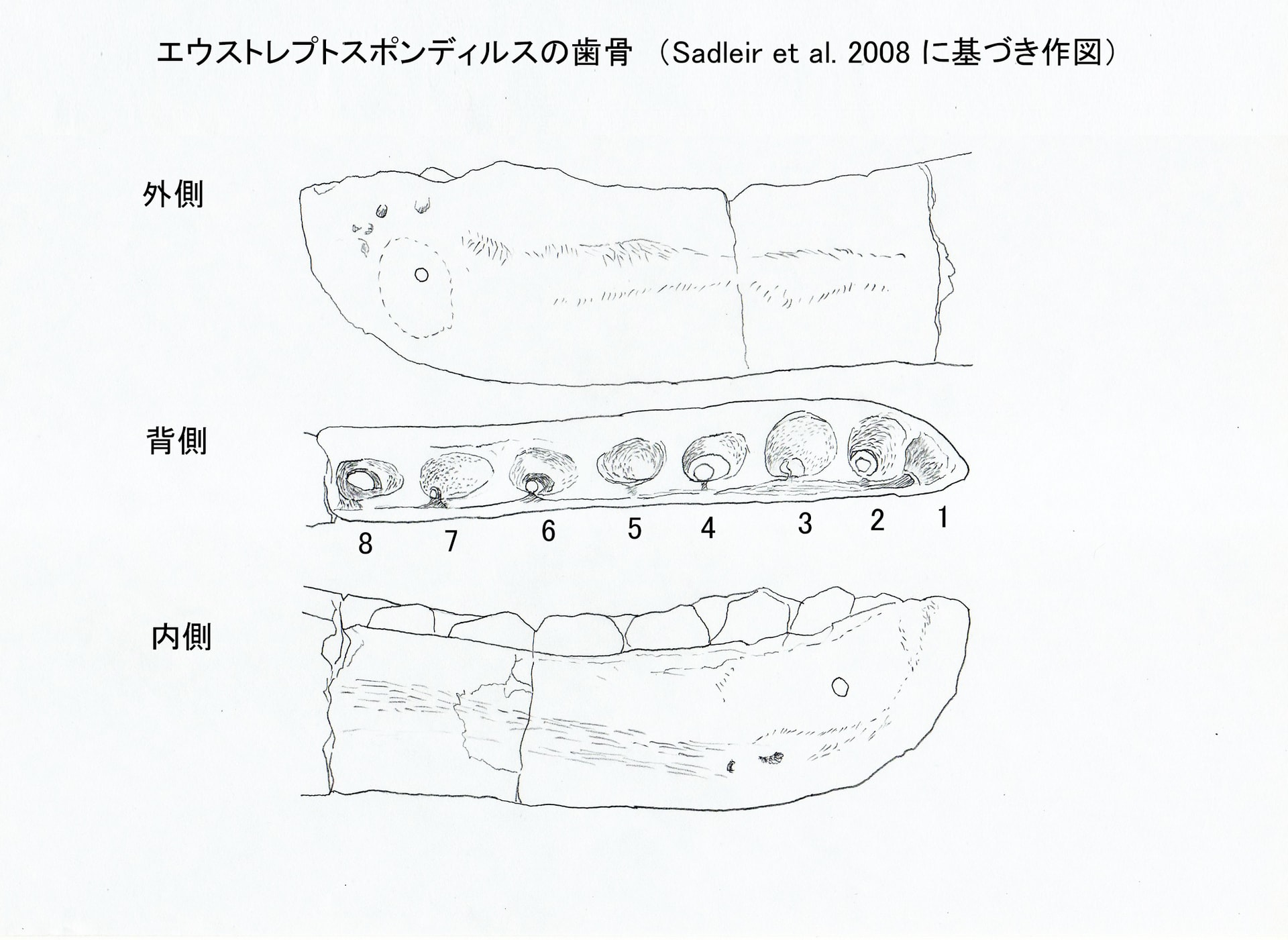
大きい画像
歯骨は、左の歯骨の前方部分と右の歯骨の後方部分が保存されていて、一部重なっているので、左右を合わせると歯骨全体の形態が得られる。左の歯骨には2, 3, 4, 6, 7, 8 番目の歯槽に置換歯が保存されている。右の歯骨には7, 8, 9, 12 番目の歯槽に置換歯がある。
側面からみると歯骨は長くスレンダーである。歯骨の前縁は緩やかにカーブしている。歯骨の腹側縁は大部分まっすぐであるが、後方では強く下降している。歯骨の背側縁は前方の25%でほぼまっすぐに延び、その後ゆるやかに下降した後、再び上昇している。そのため前方部分は少し高い位置にある。歯骨の外側面の歯列の腹側に、多数の小さな神経血管孔が分布している。これらの多くは背側縁と平行に走る浅い溝の中にある。
内側からみると歯列の内側に、一連の低い、亜三角形の歯間板がある。歯間板は歯列の中央部分で最も大きく、それより前方や後方では小さくなる。メッケル溝は前方では狭い溝であり、後方では背腹の幅が徐々に広がっている。
歯骨には少なくとも13個の歯槽がある。前方の歯槽は円形で、3番目まで大きくなっている。3番目の歯槽が歯骨の歯列の中で最も大きい。4番目の歯槽は卵形で、それより後方では歯槽は長くなり徐々に小さくなっている。歯骨の内側外側の幅は、3番目の歯槽と4番目の歯槽の間の位置で最も大きいが、それ以外の部分では一様に細い。Benson (2010)がいうようにparadental groove は前方で開いているようである。
エウストレプトスポンディルスのホロタイプ標本は全長4.5 m 、腰の高さ1.2 mと推定されるが、骨格の特徴からまだ成熟していない亜成体と考えられる。
現生のワニでは個体発生の過程で、脊椎骨の神経弓と椎体の癒合が後方から前方の順に進行する。エウストレプトスポンディルスの保存された脊椎骨の中では、第16と第21尾椎だけが完全に癒合した神経弓と椎体をもっている。前方の尾椎は、神経弓と椎体がゆるく結合しているか分離していたので、部分的に縫合線が残っていると考えられた。またより前方の脊椎でも、すべて神経弓と椎体は分離しているか、縫合線が残っていた。分離した椎体には複雑な関節面の形状が保存されていることからも、癒合してから割れたのではないことがわかる。
成熟すると仙椎は互いに癒合し、仙肋骨も含めて一体化した仙骨を形成する。エウストレプトスポンディルスの仙椎はすべて分離して見つかったことからも、未成熟であることが示唆される。また頭骨の骨についても、癒合の程度が様々であり、前頭骨と頭頂骨は癒合していたが、左右の前頭骨同士は癒合していなかった。さらに重要なことに脳函の各要素の縫合線が残っていたことから、未成熟であると考えられた。これらのことから、エウストレプトスポンディルスのホロタイプは亜成体であるが、そうすると他の基盤的テタヌラ類と異なり頸椎や胴椎に腹側のキールがないという特徴も、成長段階の影響を受けていることはないのだろうか。上腕骨などもメガロサウルスに比べてほっそりしているが、それも当然変わるだろう。いずれにしても、エウストレプトスポンディルスは未成熟のメガロサウルス類として貴重な標本にちがいない。
参考文献
Sadleir, R. W., Barrett, P. M., Powell, H. P. (2008). The anatomy and systematics of Eustreptospondylus oxoniensis, a theropod dinosaur from the Middle Jurassic of Oxfordshire, England. Palaeontographical Society Monographs 160: 1-82.
コメント ( 0 ) | Trackback ( 0 )
恐竜の類縁関係と初期進化についての新しい仮説:オルニトスケリダ
これはさすがに驚きますね。他の研究者はどうみるのか、今後の展開が楽しみです。
恐竜はまず竜盤類と鳥盤類に分かれる。Baron et al. (2017) による解析結果は、この130年もの間、揺るぎない常識であった概念を覆す仮説です。なんと鳥盤類と獣脚類が姉妹群をなし、鳥盤類+獣脚類(オルニトスケリダOrnithoscelida)が、竜脚形類とヘレラサウルス類からなる竜盤類と分岐している。恐竜は、まずオルニトスケリダと竜盤類に分かれるというわけです。ちなみに今回、エオラプトルは基盤的獣脚類に「復帰」しています。
オルニトスケリダOrnithoscelidaという名称は、1870年にHuxley がイグアノドン類、メガロサウルス類、スケリドサウルス類などを含めて命名した歴史的分類名を復活させたものです。鳥に似た後肢をもつということらしいが、今回の結果が正しければ、それが意外といい線いっていた、ということになる。
オルニトスケリダOrnithoscelidaがクレードを形成することは、21もの明確な共有派生形質で支持される。前上顎骨の鼻孔窩narial fossaの内側に孔がある、上顎骨の外側面に鋭い縦の稜がある、涙骨と上顎骨の接触により頬骨が前眼窩窓の縁から排除されている、前腹方に傾いた方形骨、・・・肩甲骨の長さが遠位の幅の3倍以上、上腕骨が近位で腹側に強く曲がる、遠位の足根骨が中足骨に癒合する、という具合である。結局、ヘテロドントサウルスのような原始的な鳥盤類と初期の獣脚類の間に、かなり共通した形質があることがポイントのようです。
なぜ今回こうなったのか?初期の恐竜進化についてのこれまでの系統解析とどこが違うのか。従来は初期の恐竜や恐竜形類の系統関係を解析する際に、獣脚類または竜盤類に重点があり、三畳紀にはまれな鳥盤類はわずかな種類しか用いられないか、上位の分類単位で代表されていた。また、竜盤類と鳥盤類に分かれるという先入観があったために、個々の形質の獲得についても多くの先験的な仮定を置いていた。今回著者らは、レソトサウルスやヘテロドントサウルス類などなるべく多くの鳥盤類を解析に含めるとともに、極力客観的に、すべての形質を適用した。従来は獣脚類や竜盤類の中で用いられていた形質を、すなおに多くの鳥盤類にも当てはめて解析した、ということのようです。過去にも何人かの研究者によって鳥盤類と獣脚類の類似性が指摘されていたが、収斂と解釈されてきた。それが今回明るみに出てきたようです。
竜盤類とオルニトスケリダの基盤的な種類(ヘレラサウルス、ヘテロドントサウルス、エオラプトルなど)からみて、恐竜の祖先はボディプランとしては小型で2足歩行性で、ものをつかむ手をもっていた。これは収斂ではなく原始状態と考えられる。
最近の研究では、初期の恐竜の食性については曖昧になってきている。パンパドロマエウス、パンファギアなどの基盤的な竜脚形類は、異歯性をもつことから早い段階で雑食性に移行したと考えられる。今回の仮説では鳥盤類と獣脚類の基盤的メンバーであるヘテロドントサウルスやエオラプトルは異歯性であり、オルニトスケリダの祖先も雑食性と考えられる。これらのことから恐竜の祖先は雑食性と解釈できる。恐竜類と最も近縁なシレサウルス類が大部分植物食性であることもこれを支持するといっている。
ヘレラサウルス類は明らかに肉食性であるが、今回の系統関係からは肉食性が派生的な形質ということになる。つまり肉食性は、ヘレラサウルス類と獣脚類で独立に進化したことになる。
さらに古生物地理学的には、シレサウルス類や最近のイギリスなどの恐竜形類の分布から、恐竜の起源はゴンドワナではなく、ローラシアで時代も少しさかのぼるといっている。
これが定説となると本当に脊椎動物学の教科書を書き換えることになる。今後どういう展開になるのか、期待されますね。
参考文献
Matthew G. Baron, David B. Norman & Paul M. Barrett (2017). A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543, 501-506. doi:10.1038/nature21700
コメント ( 0 ) | Trackback ( 0 )
アンキオルニスの体の輪郭
すでに多くの研究がなされているアンキオルニスですが、今度は骨格や羽毛ではなく、体の軟組織の外形(輪郭)を解析した仕事が報告されています。
レーザー励起蛍光法という技術がポイントです。鉱物の結晶中に含まれる有機物や無機物の不純物の組成によって、特定波長のレーザーを照射したときに異なる蛍光を発するようです。この方法を保存の良いアンキオルニスの標本に用いて、体の外形を明らかにしています。
結論として得られた体の輪郭は意外なものではないですが、これまでの骨格や羽毛の形態と、現生種との系統的な比較 extant phylogenetic bracketing からの推定を裏付けた、といっています。この復元図にはScott Hartmann が活躍していて、論文の共著者に入っています。
前肢では、肘の前方あたりにpropatagium (前飛膜、前膜)があることが四翼恐竜で初めて示されました。ここには雨覆羽の突起の痕が等間隔に並んでいるが、現生鳥類のように分化して特徴的な配列パターンをしてはいない。propatagiumは、鳥類では翼の前縁で揚力の発生に関わる機能をもっているが、アンキオルニスでも同様に滑空などの飛行機能に役立っていたかもしれない。ただし、propatagiumは明らかに飛べない平胸類やオヴィラプトロサウリア(カウディプテリクス)にもみられるそうです。
後肢では、ふくらはぎの部分が鳥類と同様にドラムスティック状をしているのが確認された。また恥骨のpubic bootの先端に軟骨らしい痕があり、さらに外側にpubic callosity (恥骨だこ)らしい組織があった。これは座った時に体を支えるのに役立つということです。
足の指の腹側には鳥類と似たfoot pad (toe pad) があり、細かい小石状の鱗で覆われていた。
尾の輪郭はスレンダーで、尾椎の形と対応している。尾の付け根の部分もわりと細めで、アンキオルニスには大きな尾大腿筋はなかったことがわかる。このことから、後肢と尾を独立して動かすことができ、飛行機能に役立ったかもしれないということです。
オープンアクセスなので全文が読めます。
参考文献
Wang, X. et al. Basal paravian functional anatomy illuminated by high-detail body outline. Nat. Commun. 8, 14576 doi: 10.1038/ncomms14576 (2017).
http://www.nature.com/articles/ncomms14576
コメント ( 0 ) | Trackback ( 0 )