獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年9月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||
| 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 21 | 22 | 23 | 24 | 25 | 26 | 27 | ||
| 28 | 29 | 30 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
フジアンヴェナトルは脚が長い
(画像は転載できないので、Natureを見てね)
フジアンヴェナトルは、後期ジュラ紀チトニアン(Nanyuan Formation)に中国福建省南平市政和県に生息したアヴィアラエ鳥類で、2023年に記載された。頭と首を除いた胴体がほぼ完全に保存されている。これは、頭骨がなく食性の決め手がないのになぜヴェナトルにしたのかな。アヴィアラエなら、オルニスとかアヴィスでいいような気がする。
他のすべての原鳥類Paravesと区別されるフジアンヴェナトルの特徴は多数の形質の組み合わせからなる。手の第I指の末節骨が他の指の末節骨よりもずっと大きい(これはアンキオルニスやアルカエオプテリクスにもみられないが、白亜紀の鳥類にはみられる)、手の指骨II-1が II-2より長い、脛骨が長く大腿骨の2倍の長さである、中足骨IIが他の中足骨よりも幅広い、などからなる。
系統解析の結果、フジアンヴェナトルはアンキオルニス、シャオティンギア、アウロルニス、エオシノプテリクスとともにアンキオルニス科に含まれた。アヴィアラエの中で最初にアンキオルニス科が分岐し、次にアルカエオプテリクスが分岐し、残りが孔子鳥類、エナンティオルニス類、真鳥類へとつながる形となった。
フジアンヴェナトルは手の指骨の比率についてはアルカエオプテリクスと似ている。アルカエオプテリクスと同様に末節骨以外の最も長い指骨はI-1である。一方シャオティンギアとアンキオルニスではII-2 である。しかし腰帯はアンキオルニスやトロオドン類の特徴を示す。短い座骨や遠位にある閉鎖孔突起はアンキオルニスと似ている。一方、幅広い恥骨エプロンpubic apronはトロオドン類と似ている。フジアンヴェナトルの後肢はいくつかの系統の形態が混在している。アルカエオプテリクスとアンキオルニスにみられるアルクトメタターサルでない足、アルカエオプテリクスと同様に中足骨IIがIIIとIVよりも幅広いこと、ドロマエオサウルス類と一部のトロオドン類にみられる蝶番関節のある中足骨IIIなどである。つまりフジアンヴェナトルにはいくつかの系統の特徴がモザイク的に混じっている。
特徴の一つは後肢の比率、つまり下肢が長いことである。著者らは前肢と後肢の骨について、代表的な多変量解析である主成分分析を行い、PC1, PC2, PC3 の三次元プロット(形態空間)の図を示している。前肢と後肢全体、または前肢の骨だけについて分析すると、フジアンヴェナトルは他のジュラ紀のアヴィアラエと近い位置にきた。ところが後肢の骨だけについて分析すると、フジアンヴェナトルはジュラ紀のアヴィアラエを含めて他のすべての獣脚類からかけ離れた位置にきた。フジアンヴェナトルは、飛翔に向けて前肢が発達するようなアヴィアラエの主流からは逸脱したものということになる。
脛骨や中足骨が長いことは、一般に高速での走行に適していることを表す。cursorial limb proportion (CLP)スコアという指標を計算して、他の獣脚類のグループと比較すると、フジアンヴェナトルは一般に疾走に適しているとされるトロオドン類やティラノサウルス類と比べても、ずっと高いスコアを示した。この数値をみるとフジアンヴェナトルは地上での高速走行に適していることになる。しかし様々な生態の現生鳥類を含めて解析してみると、フジアンヴェナトルはコウノトリやツルのような渉禽類と近い位置にきた。フジアンヴェナトルが産出した地層からは硬骨魚類、カメ、コリストデラなど水生または半水生の生物が見つかっている。フジアンヴェナトルの長い脚は沼沢地などの浅瀬に立って魚類などを捕食するためである可能性がある。残念ながら、これらを解明するために役立つ足の指骨は保存がわるく、実証するのは困難であるという。獣脚類の各グループと比較したときに、走行に適しているグループとも離れていることからは、走行とは別の機能という気もする。
参考文献
Xu, L., Wang, M., Chen, R. et al. A new avialan theropod from an emerging Jurassic terrestrial fauna. Nature 621, 336–343 (2023). https://doi.org/10.1038/s41586-023-06513-7
フジアンヴェナトルは、後期ジュラ紀チトニアン(Nanyuan Formation)に中国福建省南平市政和県に生息したアヴィアラエ鳥類で、2023年に記載された。頭と首を除いた胴体がほぼ完全に保存されている。これは、頭骨がなく食性の決め手がないのになぜヴェナトルにしたのかな。アヴィアラエなら、オルニスとかアヴィスでいいような気がする。
他のすべての原鳥類Paravesと区別されるフジアンヴェナトルの特徴は多数の形質の組み合わせからなる。手の第I指の末節骨が他の指の末節骨よりもずっと大きい(これはアンキオルニスやアルカエオプテリクスにもみられないが、白亜紀の鳥類にはみられる)、手の指骨II-1が II-2より長い、脛骨が長く大腿骨の2倍の長さである、中足骨IIが他の中足骨よりも幅広い、などからなる。
系統解析の結果、フジアンヴェナトルはアンキオルニス、シャオティンギア、アウロルニス、エオシノプテリクスとともにアンキオルニス科に含まれた。アヴィアラエの中で最初にアンキオルニス科が分岐し、次にアルカエオプテリクスが分岐し、残りが孔子鳥類、エナンティオルニス類、真鳥類へとつながる形となった。
フジアンヴェナトルは手の指骨の比率についてはアルカエオプテリクスと似ている。アルカエオプテリクスと同様に末節骨以外の最も長い指骨はI-1である。一方シャオティンギアとアンキオルニスではII-2 である。しかし腰帯はアンキオルニスやトロオドン類の特徴を示す。短い座骨や遠位にある閉鎖孔突起はアンキオルニスと似ている。一方、幅広い恥骨エプロンpubic apronはトロオドン類と似ている。フジアンヴェナトルの後肢はいくつかの系統の形態が混在している。アルカエオプテリクスとアンキオルニスにみられるアルクトメタターサルでない足、アルカエオプテリクスと同様に中足骨IIがIIIとIVよりも幅広いこと、ドロマエオサウルス類と一部のトロオドン類にみられる蝶番関節のある中足骨IIIなどである。つまりフジアンヴェナトルにはいくつかの系統の特徴がモザイク的に混じっている。
特徴の一つは後肢の比率、つまり下肢が長いことである。著者らは前肢と後肢の骨について、代表的な多変量解析である主成分分析を行い、PC1, PC2, PC3 の三次元プロット(形態空間)の図を示している。前肢と後肢全体、または前肢の骨だけについて分析すると、フジアンヴェナトルは他のジュラ紀のアヴィアラエと近い位置にきた。ところが後肢の骨だけについて分析すると、フジアンヴェナトルはジュラ紀のアヴィアラエを含めて他のすべての獣脚類からかけ離れた位置にきた。フジアンヴェナトルは、飛翔に向けて前肢が発達するようなアヴィアラエの主流からは逸脱したものということになる。
脛骨や中足骨が長いことは、一般に高速での走行に適していることを表す。cursorial limb proportion (CLP)スコアという指標を計算して、他の獣脚類のグループと比較すると、フジアンヴェナトルは一般に疾走に適しているとされるトロオドン類やティラノサウルス類と比べても、ずっと高いスコアを示した。この数値をみるとフジアンヴェナトルは地上での高速走行に適していることになる。しかし様々な生態の現生鳥類を含めて解析してみると、フジアンヴェナトルはコウノトリやツルのような渉禽類と近い位置にきた。フジアンヴェナトルが産出した地層からは硬骨魚類、カメ、コリストデラなど水生または半水生の生物が見つかっている。フジアンヴェナトルの長い脚は沼沢地などの浅瀬に立って魚類などを捕食するためである可能性がある。残念ながら、これらを解明するために役立つ足の指骨は保存がわるく、実証するのは困難であるという。獣脚類の各グループと比較したときに、走行に適しているグループとも離れていることからは、走行とは別の機能という気もする。
参考文献
Xu, L., Wang, M., Chen, R. et al. A new avialan theropod from an emerging Jurassic terrestrial fauna. Nature 621, 336–343 (2023). https://doi.org/10.1038/s41586-023-06513-7
コメント ( 0 ) | Trackback ( 0 )
バラウル

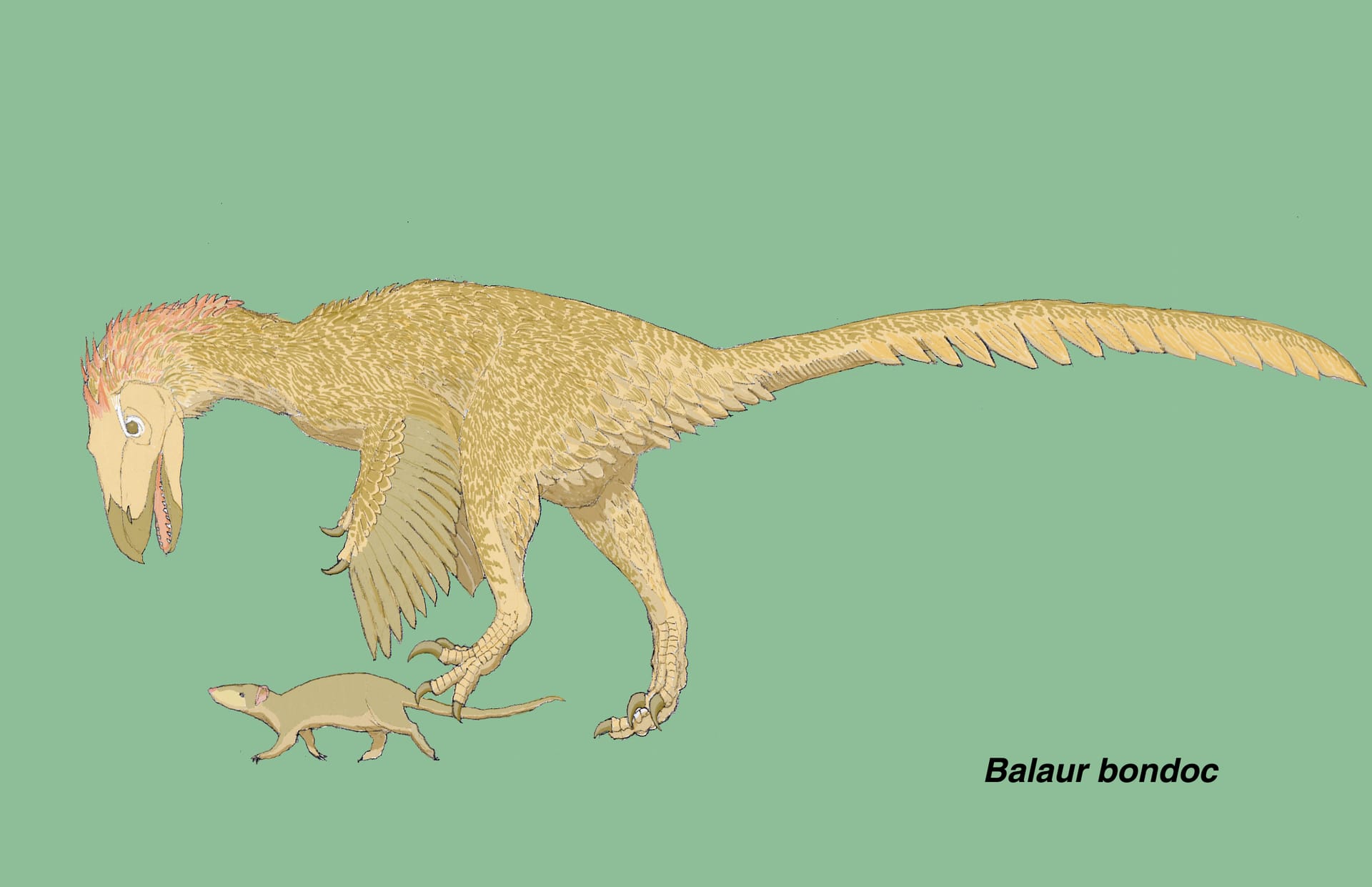
大きい画像
恐竜研究の進展は本当に日進月歩で、油断もすきもないという話である。
バラウル・ボンドクは、白亜紀後期マーストリヒト期のルーマニア(ハチェグ島)から発見された奇妙な獣脚類で、白亜紀後期のヨーロッパでは最も完全な獣脚類とされた。ホロタイプは同一個体の胴体の部分骨格で、胴椎、仙椎、尾椎と胸帯、前肢、腰帯、後肢を含んでいる。最初の記載 (Csiki et al., 2010) 、ドロマエオサウルス類の総説 (Turner, Makovicky & Norell, 2012)、 バラウルのフル記載論文 (Brusatte et al., 2013) では、系統解析の結果、ヴェロキラプトルに最も近縁なドロマエオサウルス類と結論されていた。第2指の他に第1指にも大きな鎌状のカギ爪をもつ、二重の鎌double-sickleのドロマエオサウルス類として注目された。なにしろBrusatte et al. (2013) の論文には、ヴェロキラプトルそっくりのシルエットが載っているので、こぞってイラストを描いた恐竜ファンも多いはずだ。私もこのフル記載が決定版と思ってすっかり安心していた。ところが、全部読まないうちに風向きが変わってきた。
研究の結果、バラウルは他のドロマエオサウルス類やほとんどの非鳥型獣脚類にはみられない、一連の固有形質をもつことがわかってきた。たとえば手根骨と中手骨が癒合していること、手の第3指が退化していること、足根骨と中足骨が癒合していることなどである。ドロマエオサウルス類としては、バラウルは非常に奇妙で独特の特徴をもっていることになる。ところが、これらの多くは、原始鳥類であるアヴィアラエ類にはよくみられるものでもある。
その後Godefroit et al. (2013a) (アウロルニスの論文)はパラヴェス類の系統解析にバラウルを含めた結果、バラウルはアルカエオプテリクスより派生的なアヴィアラエ類に位置づけられた。それとは独立にFoth, Tischlinger &Rauhut (2014) (アルカエオプテリクスの論文)が行った系統解析でも、多少の違いはあるが、やはりアルカエオプテリクスより派生的なアヴィアラエ類となった。そこで、Cau et al. (2015) はバラウルの系統上の位置を主題とした研究を行い、2つのデータセットを用いて新たに解析を行った。その結果、やはりアヴィアラエ類という位置づけが支持された。Cau et al. (2015) によると、バラウルがドロマエオサウルス類であるという可能性が完全に否定されたわけではないが、当面はアヴィアラエ類という枠組みで考えるべきである、という。
実際の分岐分析は、860とか1500もの形質について計算しているので1つや2つの形質が決め手になるというものではないだろう。しかし具体的に個々の骨がどんな形態なのか見ないと、イメージがわかない。著者らは系統関係に重要と思われる多くの形質について解説している。バラウルが、ドロマエオサウルス類とアヴィアラエ類のどちらに近縁なのかという観点から見ていくと面白い。

Copyright 2015 Cau et al.
手については、A: バラウル、B: ジョウオルニス、C: サペオルニス、D: デイノニクスを並べて比較している。
バラウルの手では、遠位の手根骨が中手骨の近位端と癒合している。これはドロマエオサウルス類には全くみられない。手根骨と中手骨の癒合は、アヴィミムスやモノニクスのような2、3の非鳥型獣脚類と、パイゴスティル類にみられる。特に、バラウルにみられる骨の癒合のパターンは、最も基盤的なパイゴスティル類(コンフキウソルニス、シノルニス、サペオルニス、ジョウオルニスなど)と共通しているという。
バラウルでは、半月形の手根骨(lsc、中手骨と癒合している)が第 II 中手骨と第 III 中手骨の根元にある。また第 I 中手骨の近位端は遠位端より細くなっていて、中手骨の内側縁が斜めになっている。ほとんどの非鳥型獣脚類では、半月形の手根骨(右のデイノニクスのusc)は第 I 中手骨と第 II 中手骨にまたがっている。また第 I 中手骨の近位端は細くなっていない。つまりバラウルでは半月形の手根骨の位置が、側方にずれている。これはパイゴスティル類(コンフキウソルニス、シノルニス、サペオルニスなど)と似ている。またパイゴスティル類はバラウルと同様に、第 I 中手骨の近位端が細くなっていて、内側縁が斜めになっている。
さらに第III指の状態が特徴的である。バラウルでは第 II 中手骨の側面に稜があり、遠位で第 III 中手骨と接しているので、第 II 中手骨と第 III 中手骨の間のスペースが閉じている。基盤的アヴィアラエ類では、サペオルニスのように第 II 中手骨と第 III 中手骨がまっすぐで密着しているものや、バラウルに似て遠位で接しているもの、遠位端が完全に癒合しているものなど様々である。中手骨の間のスペースが閉じている状態は、コンフキウソルニス、ジェホロルニス、ジシャンゴルニス、エナンティオルニスなどにみられる。
バラウルの第 III 中手骨は遠位端が単純な形で、はっきりした関節顆になっていない。ドロマエオサウルス類はほとんどの非鳥型獣脚類と同様に、よく発達した関節顆をもっている。バラウルの状態は、ティラノサウルス類以外では基盤的パイゴスティル類と派生的な鳥類に似ている。
バラウルの第 III 指は極端に縮小しており、末節骨を含めて遠位の指骨が欠けている。バラウルの第 III 指の唯一の指骨は先細りで、遠位端に小さな関節面があるので、もう1個非常に小さい指骨があったかもしれない。それでも2個である。このような縮小はドロマエオサウルス類(指骨は4個)にはみられないが、シノルニス、サペオルニスなどのパイゴスティル類には普通にみられる(指骨が2個以下)。
バラウルでは腰帯の骨が完全に癒合coossificationしており、腸骨/恥骨間および腸骨/座骨間の縫合線が閉じている。最も基盤的なアヴィアラエ類を含めて、ほとんどのテタヌラ類では腰帯の骨が完全には癒合していない。一方、ケラトサウルス類、一部のコエルロサウルス類(アヴィミムス)、鳥胸類ornithothoracines(アプサラヴィス、パタゴプテリクス、シノルニスなど)では腰帯の骨が完全に癒合している。
バラウルでは、左右の恥骨が側方に膨らんで、腹側で急に狭まっているので、骨盤腔pelvic canalが広くなっている(VでなくU字形)。このような状態は、ヴェロキラプトルやバンビラプトルなどを含めてほとんどの獣脚類と異なっている。バラウルのようなU字形の骨盤は、パイゴスティル類(コンコルニス、サペオルニスなど)にみられる。

Copyright 2015 Cau et al.
足については、A: ヴェロキラプトル、B: バラウル、C: ジョウオルニスを並べている。
バラウルでは、脛骨の遠位端と近位の足根骨が癒合して脛足根骨tibiotarsusを形成している。このような癒合は、コエルロサウルス類の中ではモノニクスのようなアルバレッツサウルス類やアヴィミムスのような一部のオヴィラプトロサウリアにみられる。またアヴィアラエ類の中では、完全に癒合した脛足根骨はアルカエオプテリクスよりも派生的な種類(アプサラヴィス、コンフキウソルニス)にみられる。
バラウルでは、足根中足骨tarsometatarsus部分の骨が広範に癒合している。ヴェロキラプトルのようなほとんどの非鳥型獣脚類では、このような癒合はみられない。遠位の足根骨と中足骨の近位端の癒合は多くのマニラプトル類にみられるが、中足骨同士の広範な癒合はバラウルとパイゴスティル類にしかみられない。
バラウルの足で最も特徴的なのが第 I 指だろう。バラウルの足の第 I 指は他の指に比べて縮小していない。ドロマエオサウルス類を含めてほとんどの非鳥型獣脚類では、第 I 指のカギ爪は相対的に小さい。一方、バラウルのように他の指に比べて縮小していない、大きな鎌状の第 I 指のカギ爪は、多くの基盤的アヴィアラエ類にもみられる(コンフキウソルニス、ジシャンゴルニス、パタゴプテリクス、サペオルニス、ジョウオルニス)。また、バラウルの第 I 指の第1指骨は 第II , III, IV指の第1指骨と同じくらいの長さであるが、これも基盤的アヴィアラエ類と同様である。さらに、よく発達した関節面からバラウルの第 I 指は完全に機能的と思われるが、これも鳥類にはみられるが非鳥型獣脚類にはみられないものである。なるほど、第 II 指の他に第 I 指も大きいのは鳥ならば普通のことというわけである。
バラウルの第 II 指のカギ爪は、多くのデイノニコサウリアと同様に第III、第IV指のカギ爪よりも大きく肥大している。しかし、Brusatte et al. (2013)は、バラウルの第 II 指のカギ爪は、多くのドロマエオサウルス類にみられるような顕著な鎌状の形や屈筋結節を示していないと記している。バラウルの状態と似た、大きくほどほどにカーブしたカギ爪のあるがっしりした第 II 指は、いくつかのアヴィアラエ類にみられるという。
ドロマエオサウルス類を含めてほとんどの獣脚類では、足の第III 指の末節骨より一つ手前の指骨(第3指骨)がその手前の指骨と同じくらいか、より短い。ところがバラウルでは、この指骨が比較的長く、手前の指骨の1.2倍の長さがある。この状態は多くのアヴィアラエ類にみられるのと同様であるという(コンコルニス、サペオルニス、ジョウオルニス)。
Cau et al. (2015) は、手が機能的な第III 指を欠くこと、足の第 II 指のカギ爪を使うための第II中足骨の関節面が蝶番形でないこと、足の第 II 指のカギ爪が他のドロマエオサウルス類ほど鎌状にカーブしていないことから、バラウルは雑食性ないし植物食の可能性が高いといっている。このことはアヴィアラエ類という系統的位置とも一致するという。骨盤腔が広いのも長い腸を収めることと対応するといっている。それでは、「ドロマエオ復元」に比べて「アヴィアラエ復元」はガラリと変わったのかというと、そうでもない。Cau et al. (2015) の論文も全身復元図を載せているが、第 II 指のカギ爪をあまり持ち上げていない、「おとなしいヴェロキラプトル」みたいになっている。
仕方が無いので小型で樹上性・雑食性の祖先が地上に降りて大型化した動物としよう。しかしアヴィアラエ類としても異様な動物である。肉食傾向の強い雑食性で、普段は昆虫や果実などを食べるが、機会があればトカゲなどの小動物も捕らえるということでどうだろう。
参考文献
Brusatte SL, Vremir M, Csiki-Sava Z, Turner AH, Watanabe A, Erickson GM, Norell MA. (2013). The Osteology of Balaur bondoc, an island-dwelling dromaeosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Romania. Bulletin of the American Museum of Natural History 374:1-100
Andrea Cau, Tom Brougham and Darren Naish (2015), The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): dromaeosaurid or flightless bird? PeerJ 3:e1032; DOI 10.7717/peerj.1032
コメント ( 0 ) | Trackback ( 0 )
アウロルニス
アウロルニスは、ジュラ紀中期から後期(Tiaojishan Formation)に中国遼寧省に生息した原始的なアヴィアラエ鳥類で、最近Godefroit et al. (2013)によって記載された。Natureの論文の共著者の一人、Francois Escuillie とは、ミネラルフェアで化石を販売している陽気なフランス人、Eldoniaのフランソワさんである。すごいですね。こないだ「This is our work.」といって宣伝していたので読んだのであるが、予習していたらもっと詳しい話が聞けたのに残念である。
最近のジュラ紀中期から後期の遼寧省からの、鳥類に近縁な小型獣脚類の発見は、鳥類の祖先としての始祖鳥アルカエオプテリクスの地位を脅かしつつある。もしアルカエオプテリクスが鳥類の祖先ではなく、デイノニコサウリアに属するとすれば、前肢だけで羽ばたくという典型的な鳥類の飛翔が、独立に2回生じたか、あるいは後にデイノニコサウリアで失われたことになる。
今回Godefroit et al. (2013) は遼寧省で発見されたアウロルニスの全身骨格を記載し、これを包括的な系統解析に含めることにより、次のことを示唆した。1)系統解析の結果、アウロルニスは最も基盤的なアヴィアラエとされた。2)アルカエオプテリクスはアヴィアラエに属することが確認された。3)この解析ではトロオドン類がアヴィアラエの姉妹群として位置付けられた。4)パラヴェスParavesの中で、飛翔powered flight は一回だけ生じたことが支持された。5)ジュラ紀中期から後期の頃には、アヴィアラエとパラヴェスの初期の多様化が既に起こっていた。
アウロルニスに特徴的な形質は、手の第1指の第1指骨が撓骨よりも顕著に太い、腸骨の後寛骨臼突起postacetabular processが腹側にはっきり屈曲しておらず、背縁が水平である、座骨の遠位端が背腹に拡張し、カギ状の腹側突起とより長い背側遠位突起からなる、第1中足骨が華奢で細長い(第3中足骨の約30%)、であるという。
この系統解析の結果からみると、基盤的なアヴィアラエ(アウロルニス、アンキオルニス、シャオティンギア)は、ジュラ紀中期から後期の中国北部では既に多様化していたことがわかる。ドイツのアルカエオプテリクスを含めるとジュラ紀末には、アヴィアラエ鳥類はユーラシアに広く分布していたことになる。一方、ドロマエオサウルス類とトロオドン類はジュラ紀のアジアからは見つかっておらず、ジュラ紀後期のヨーロッパからの分離した歯が暫定的にドロマエオサウルス類とされているに過ぎない。熱河生物群の化石から、ドロマエオサウルス類とトロオドン類は、白亜紀に入って急速に発展・多様化したことになる。
参考文献
Pascal Godefroit, Andrea Cau, Hu Dong-Yu, Francois Escuillie, Wu Wenhao & Gareth Dyke (2013) A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds. Nature 498, 359-362.
最近のジュラ紀中期から後期の遼寧省からの、鳥類に近縁な小型獣脚類の発見は、鳥類の祖先としての始祖鳥アルカエオプテリクスの地位を脅かしつつある。もしアルカエオプテリクスが鳥類の祖先ではなく、デイノニコサウリアに属するとすれば、前肢だけで羽ばたくという典型的な鳥類の飛翔が、独立に2回生じたか、あるいは後にデイノニコサウリアで失われたことになる。
今回Godefroit et al. (2013) は遼寧省で発見されたアウロルニスの全身骨格を記載し、これを包括的な系統解析に含めることにより、次のことを示唆した。1)系統解析の結果、アウロルニスは最も基盤的なアヴィアラエとされた。2)アルカエオプテリクスはアヴィアラエに属することが確認された。3)この解析ではトロオドン類がアヴィアラエの姉妹群として位置付けられた。4)パラヴェスParavesの中で、飛翔powered flight は一回だけ生じたことが支持された。5)ジュラ紀中期から後期の頃には、アヴィアラエとパラヴェスの初期の多様化が既に起こっていた。
アウロルニスに特徴的な形質は、手の第1指の第1指骨が撓骨よりも顕著に太い、腸骨の後寛骨臼突起postacetabular processが腹側にはっきり屈曲しておらず、背縁が水平である、座骨の遠位端が背腹に拡張し、カギ状の腹側突起とより長い背側遠位突起からなる、第1中足骨が華奢で細長い(第3中足骨の約30%)、であるという。
この系統解析の結果からみると、基盤的なアヴィアラエ(アウロルニス、アンキオルニス、シャオティンギア)は、ジュラ紀中期から後期の中国北部では既に多様化していたことがわかる。ドイツのアルカエオプテリクスを含めるとジュラ紀末には、アヴィアラエ鳥類はユーラシアに広く分布していたことになる。一方、ドロマエオサウルス類とトロオドン類はジュラ紀のアジアからは見つかっておらず、ジュラ紀後期のヨーロッパからの分離した歯が暫定的にドロマエオサウルス類とされているに過ぎない。熱河生物群の化石から、ドロマエオサウルス類とトロオドン類は、白亜紀に入って急速に発展・多様化したことになる。
参考文献
Pascal Godefroit, Andrea Cau, Hu Dong-Yu, Francois Escuillie, Wu Wenhao & Gareth Dyke (2013) A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds. Nature 498, 359-362.
コメント ( 0 ) | Trackback ( 0 )
エピデクシプテリクス1

エピデクシプテリクスは、ジュラ紀中期から後期の中国内モンゴル自治区に生息した奇妙な鳥類(鳥群、アヴィアラエ)の一員で、最近(2008年)Natureに報告された。進化した鳥類の形質と、オヴィラプトロサウリアなど他の獣脚類の系統にみられる形質が奇妙に混じっており、系統解析の結果からエピデンドロサウルスと姉妹群をなすとされている。ハトくらいの大きさで、尾の先端に2対の長いリボン状の尾羽をもっていたが、四肢には翼の痕跡はないという。
頭骨は、オヴィラプトロサウルス類のように短く丈が高い。外鼻孔は吻の上方にあり、前方の歯は上顎・下顎とも、マシアカサウルスのように強く傾斜している。また、前方の歯は後方の歯よりもずっと大きいが、これは基盤的なオヴィラプトロサウルス類や基盤的なテリジノサウロイドと同様である。頚椎は9個、胸椎は14個あり、合仙骨は7個の椎骨からなる。尾椎のうち前方の6個は短く幅広く、後方の10個は原始的な尾端骨に似た構造をなすが、互いに癒合してはいないという。
肩甲骨は他の進化したマニラプトル類と同様に上腕骨より短く、烏口骨は長方形に近い。上腕骨は大腿骨と同じくらいの長さで、これは他の多くの原始的なアヴィアラエより長い。尺骨は後方に反っており、手は細長い。腰帯のうち、腸骨のpreacetabular processが長く、その前縁が強く凸型であるが、これは進化した鳥類と同様である。また獣脚類には珍しく、恥骨が座骨より短い。恥骨は前腹方を向いており、pubic bootはない。座骨は後方に曲がっており、扁平で、遠位端に向かうにつれて幅広くなっている。
系統解析の結果、エピデクシプテリクスとエピデンドロサウルスは、アヴィアラエの基盤的な位置に、単系のスカンソリオプテリクス科を構成すると考えられた。エピデクシプテリクスとエピデンドロサウルスは、鳥類にみられる多くの派生的な形質(上腕骨が大腿骨と同じくらい長い、腸骨のpreacetabular processが長く、その前縁が強く凸型など)を示す一方で、オヴィラプトロサウルス類やテリジノサウロイドと顕著な類似性を示す。例えば、短く丈の高い頭骨、外鼻孔の位置が高い、前方の歯が大きい、前端が下方に曲がり、背側が強くふくらんだ下顎、大きな外側下顎窓などである。また恥骨が座骨より短いなどの腰帯の特徴は、獣脚類の中でも独特であるという。
鳥になってしまうとあまり興味はわかないが、この動物は変わっている。歯のある鳥類も珍しくないとはいえ、これは歯は大きいし翼はないし、鳥らしくない。頭骨の特殊化は食性と関連しているかもしれないが、昆虫食なのか、植物の実でもとったのであろうか。
参考文献
Zhang, F., Zhou, Z., Xu, X., Wang, X. and Sullivan, C. 2008. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature 455, 1105-1108.
コメント ( 2 ) | Trackback ( 0 )