獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2016年2月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||
| 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 21 | 22 | 23 | 24 | 25 | 26 | 27 | ||
| 28 | 29 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
フクイベナトル
フクイベナトル (フクイベナートル、フクイヴェナトル)
ざ、雑食‥‥‥orz ペーパークラフトも買ったのに、ショック大きい‥‥。
基本情報は「恐竜の楽園」見て下さい。(やる気がない)Scientific Reports は無料で読めるので、論文の図を見て下さい。(終了)
これは、こんなビザールな獣脚類だったんですね。そもそも歯が。。。
基盤的なマニラプトラの一種で、コンプソグナトゥス科やオルニトミモサウリア、オルニトレステス、より派生的なグループ(テリジノなど)とポリトミーをなす。この辺りで肉食でがんばっているのはオルニトレステスくらいで、みな雑食・植物食への道を歩み始めたころなので、フクイベナトルはそのプロトタイプ的な一種ということか。
他のどの獣脚類にもみられない独特の特徴が14もあるのですが、私がわかりやすいと思うのは、「高度に異歯性の歯列」ですね。典型的な獣脚類の歯と異なり、すべての歯で前縁にも後縁にも鋸歯がなく、断面が卵形で、また位置によって大きさと形が異なっている。前上顎骨の歯は分離したものが2本あるが、形が違っている。最前方の歯?は小さく、先がスプーン型spatulateで、ある程度インキシヴォサウルスの歯と似ている。もう1本の歯は側面から見ると葉状leaf-likeで、前方に稜縁がある。前方の上顎骨歯は長く、後方にカーブしている。一方、後方の上顎骨歯と歯骨歯は葉状で、前後に対称形symmetricalである。
あと重要らしい特徴が、頚椎に複雑なlamina system(神経弓の突起をつなぐ板状の稜など)が発達していることで、靭帯のための溝をなすとあるので、首が長くなることに関係しているようですね。10個の頚椎が保存されていて、環椎はないので、フクイベナトルは少なくとも11個の頚椎をもっていた。これは他のほとんどの獣脚類よりも多い。また前方と中央の頚椎で、神経棘が2分岐しているのは他のどの獣脚類にもみられず、一部の竜脚類にしかみられない。
「恐竜の楽園」で指摘されている通り、骨格図のスケールがおかしいようです。また、本文ではpromaxillary fenestra なのに、Diagnosis とFig.2 ではpremaxillary fenestra になっている。本文では10個の頚椎とあるのに、Holotypeの説明では8個の頚椎となっている。
中央の尾椎の前関節突起の先端が2分岐しているなど、固有の形質の説明はあるが、前関節突起や血道弓の長さがドロマエオサウルス類と比べてどうとかいう解説はないです。一方、足の第2指の末節骨は欠けているようですが、指骨II-1 の形と末節骨が大きい点でデイノニコサウリアと似ているとある。
結局どういう点がドロマエオサウルス類と似ていて、どういう点が違うのか、ということが知りたいですよね。
前上顎骨、上顎骨、涙骨、前頭骨、鱗状骨、傍後頭骨突起などにはドロマエオサウルス類と似た特徴がある。一方で、烏口骨、尺骨、中手骨、大腿骨の小転子・大転子、坐骨などには多数の原始的な特徴がある。
このようにモザイク的な特徴を示すフクイベナトルを入れて分岐分析すると、コエルロサウリアの系統に大きな影響があり、いくつものクレードが崩壊してしまったそうです。ドロマエオサウルス科自体が崩壊しています。いろいろとやっかいな種のようです。
日本初のドロマエオサウルス類の全身骨格!とはならなかったが、コエルロサウリアの系統に与えるインパクトは重大で、その意味では重要な発見であると、ポジティブにとらえることができるはずだ。あれ、なにか自分に言い聞かせている感が。。。
参考文献
Azuma, Y. et al. A bizarre theropod from the Early Cretaceous of Japan highlighting mosaic evolution among coelurosaurians. Sci. Rep. 6, 20478; doi: 10.1038/srep20478 (2016).
ざ、雑食‥‥‥orz ペーパークラフトも買ったのに、ショック大きい‥‥。
基本情報は「恐竜の楽園」見て下さい。(やる気がない)Scientific Reports は無料で読めるので、論文の図を見て下さい。(終了)
これは、こんなビザールな獣脚類だったんですね。そもそも歯が。。。
基盤的なマニラプトラの一種で、コンプソグナトゥス科やオルニトミモサウリア、オルニトレステス、より派生的なグループ(テリジノなど)とポリトミーをなす。この辺りで肉食でがんばっているのはオルニトレステスくらいで、みな雑食・植物食への道を歩み始めたころなので、フクイベナトルはそのプロトタイプ的な一種ということか。
他のどの獣脚類にもみられない独特の特徴が14もあるのですが、私がわかりやすいと思うのは、「高度に異歯性の歯列」ですね。典型的な獣脚類の歯と異なり、すべての歯で前縁にも後縁にも鋸歯がなく、断面が卵形で、また位置によって大きさと形が異なっている。前上顎骨の歯は分離したものが2本あるが、形が違っている。最前方の歯?は小さく、先がスプーン型spatulateで、ある程度インキシヴォサウルスの歯と似ている。もう1本の歯は側面から見ると葉状leaf-likeで、前方に稜縁がある。前方の上顎骨歯は長く、後方にカーブしている。一方、後方の上顎骨歯と歯骨歯は葉状で、前後に対称形symmetricalである。
あと重要らしい特徴が、頚椎に複雑なlamina system(神経弓の突起をつなぐ板状の稜など)が発達していることで、靭帯のための溝をなすとあるので、首が長くなることに関係しているようですね。10個の頚椎が保存されていて、環椎はないので、フクイベナトルは少なくとも11個の頚椎をもっていた。これは他のほとんどの獣脚類よりも多い。また前方と中央の頚椎で、神経棘が2分岐しているのは他のどの獣脚類にもみられず、一部の竜脚類にしかみられない。
「恐竜の楽園」で指摘されている通り、骨格図のスケールがおかしいようです。また、本文ではpromaxillary fenestra なのに、Diagnosis とFig.2 ではpremaxillary fenestra になっている。本文では10個の頚椎とあるのに、Holotypeの説明では8個の頚椎となっている。
中央の尾椎の前関節突起の先端が2分岐しているなど、固有の形質の説明はあるが、前関節突起や血道弓の長さがドロマエオサウルス類と比べてどうとかいう解説はないです。一方、足の第2指の末節骨は欠けているようですが、指骨II-1 の形と末節骨が大きい点でデイノニコサウリアと似ているとある。
結局どういう点がドロマエオサウルス類と似ていて、どういう点が違うのか、ということが知りたいですよね。
前上顎骨、上顎骨、涙骨、前頭骨、鱗状骨、傍後頭骨突起などにはドロマエオサウルス類と似た特徴がある。一方で、烏口骨、尺骨、中手骨、大腿骨の小転子・大転子、坐骨などには多数の原始的な特徴がある。
このようにモザイク的な特徴を示すフクイベナトルを入れて分岐分析すると、コエルロサウリアの系統に大きな影響があり、いくつものクレードが崩壊してしまったそうです。ドロマエオサウルス科自体が崩壊しています。いろいろとやっかいな種のようです。
日本初のドロマエオサウルス類の全身骨格!とはならなかったが、コエルロサウリアの系統に与えるインパクトは重大で、その意味では重要な発見であると、ポジティブにとらえることができるはずだ。あれ、なにか自分に言い聞かせている感が。。。
参考文献
Azuma, Y. et al. A bizarre theropod from the Early Cretaceous of Japan highlighting mosaic evolution among coelurosaurians. Sci. Rep. 6, 20478; doi: 10.1038/srep20478 (2016).
コメント ( 2 ) | Trackback ( 0 )
ドロマエオサウルス補足
ドロマエオサウルスの前上顎骨歯について、前の記事中の文言を一部訂正し、ここに補足説明します。
ヴェロキラプトルでは2番目の歯が最も大きいのに対して、ドロマエオサウルスでは4本とも同じくらいとされる理由は、分離した前上顎骨歯が見つかっているため。
Colbert and Russell (1969) によると、歯槽に保存されているのは左の前上顎骨の後半部分の2本(3番と4番)だけであるが、その他に4本の分離した前上顎骨歯が見つかっている。形態から、これらのうち2本は右側、2本は左側のものである。さらに前縁と後縁の近接の程度(D字形に近い)から、右の1本と左の1本は1番目の前上顎骨歯と考えられる。よって左側の残りの1本は2番目となり、左側は一応、1、2、3、4が揃っていることになる。そのためCurrie (1995)の論文中で、大きさが4本とも同じくらいとされている。それなりのロジックがあって結論されているのが面白い。
ドロマエオサウルスの最大の特徴である、歯の前縁のねじれを確認しようと思って、Colbert and Russell (1969)を参照したのであるが、なんと前上顎骨歯の写真はあるが、上顎骨歯と歯骨歯の写真はないようだ。上顎骨歯については、文章で記述されている。
参考文献
Colbert, E.H. and Russel, D.A. (1969). The small cretaceous dinosaur Dromaeosaurus. American Museum Novitates 2380: 1-49.
コメント ( 0 ) | Trackback ( 0 )

ドロマエオサウルス


大きい画像
ドロマエオサウルスは、白亜紀後期カンパニア期(Oldman Formation)にカナダのアルバータ州に生息した中型のドロマエオサウルス類で、Matthew and Brown (1922) によって記載された。Matthew and Brown (1922)の記載は、部分的にクリーニングされた標本に基づく予備的なものだったが、その後Colbert and Russell (1969) によってより完全な記載がなされた。さらにCurrie (1995)は、ホロタイプ標本の再クリーニングとCTスキャンによって、頭蓋と脳函の形態について新しい情報を追加し、頭骨の復元を行った。
ホロタイプAMNH FARB 5356 には、部分的な頭骨と下顎、舌骨、足の骨、左の第1中手骨が含まれている。
ホロタイプの頭骨は、上顎、下顎、後頭部の骨が主で、頭蓋天井の大部分は見つかっていない。つまり鼻骨、上顎骨の上部、涙骨のほとんど、後眼窩骨などは保存されていない。前頭骨と脳函はある。

大きい画像
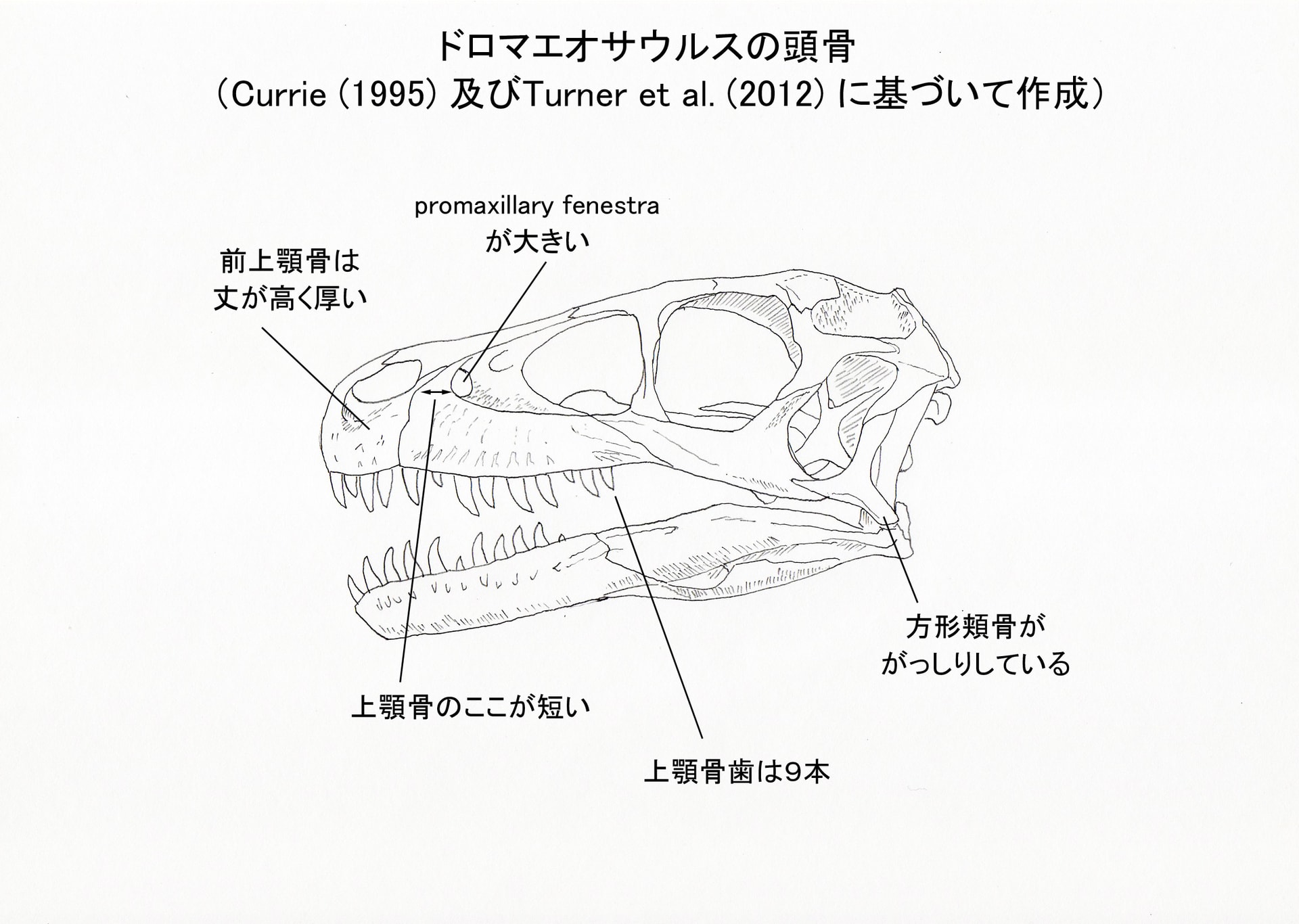
Turner et al. (2012) によると、ドロマエオサウルスは以下の形質の組み合わせと固有形質により識別される中型のドロマエオサウルス類であるという(これらの大部分はCurrie (1995) からそのまま採用している)。ここでは主なものを骨ごとにまとめてみる。
歯:9本の上顎骨歯;上顎骨歯と歯骨歯の前縁anterior carinaが(先端付近では正中にあるが)舌側へねじれている;前縁の鋸歯と後縁の鋸歯の大きさがほぼ同じくらいである。
前上顎骨:(他のドロマエオサウルス類よりも)前上顎骨の丈が高く厚い。
上顎骨:上顎骨の前眼窩窩より前方の部分が前後に短い;拡大したpromaxillary fenestra (または極端に前腹方にあるmaxillary fenestra)。
方形頬骨:方形頬骨はがっしりしているstout。
前頭骨:前頭骨の先端が扁平である;前頭骨の後眼窩骨突起と眼窩の背側縁の境界がはっきりしている;鼻骨と前頭骨の間の縫合線がV字形;前頭骨の前方に涙骨と関節するための深い切れ込みnotchがある。
その他、口蓋骨の形質が1つ、脳函の形質が3つある。ドロマエオサウルスは他のドロマエオサウルス類と異なり、脳函がほとんど含気化していないという。


大きい画像
前上顎骨は、2本の歯のついた後半部分だけが保存されているが、他のドロマエオサウルス類と同様に4本の歯があったと考えられる。ヴェロキラプトルなどでは前上顎骨の2番目の歯が長さ・直径とも最も大きいが、ドロマエオサウルスでは4本とも同じくらいの大きさと考えられる(分離した前上顎骨歯が見つかっている)。
左の上顎骨に、9個の歯槽がある。デイノニクスでは15個、ヴェロキラプトルでは10個、サウロルニトレステスでは少なくとも10個あるという。
涙骨はほとんど失われているが、右の頬骨に付着した小さな断片と、いくつかの分離した断片がある。その形状から、ドロマエオサウルスにはデイノニクスなどにみられるような、眼窩の前背方で側方に突出した膨らみがあったと思われるという。
方形頬骨はT字形で、ヴェロキラプトルなどと似ているが、デイノニクス、ヴェロキラプトル、サウロルニトレステスのものよりもがっしりしている。鱗状骨は、顕著に側方にのびた腹側方突起ventrolateral process をもつなど、ドロマエオサウルス類の特徴を示している。
Turner et al. (2012) の論文中にもドロマエオサウルスの頭骨復元図があるが、ちょっと長めのツァーガン的な顔になっている。これをよくみると疑問な点がある。「上顎骨の前眼窩窩より前方の部分が前後に短い」はCurrie (1995) にはないので、Turnerらが追加した特徴と思われるが、この通りになっていない。図では前眼窩窩より前方の部分が妙に長くなっている。この点で、本文の記述と頭骨復元図が一致していない。Currie (1995)の頭骨復元図の方が信頼できるように思われた。
Turner et al. (2012) の系統解析では、ドロマエオサウルス、ユタラプトル、アキロバトル、アトロキラプトルがドロマエオサウルス亜科Dromaeosaurinae に含まれた。
ヴェロキラプトルなどと比べると、ドロマエオサウルス亜科はどれも不完全である。ユタラプトルやアキロバトルの姿を描こうとすればドロマエオサウルスが参考になるはずだが、ドロマエオサウルス自体が不完全という感じである。ヴェロキラプトルなどよりも上顎、下顎ががっしりしているので、より大型の獲物に挑みそうな、獰猛な印象はある。現代でも生存できそうな洗練されたハンターと考えたい。
参考文献
Currie, P.J. (1995). New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal of Vertebrate Paleontology 15: 576-591.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )
アダサウルスの特徴と系統的位置

大きい画像
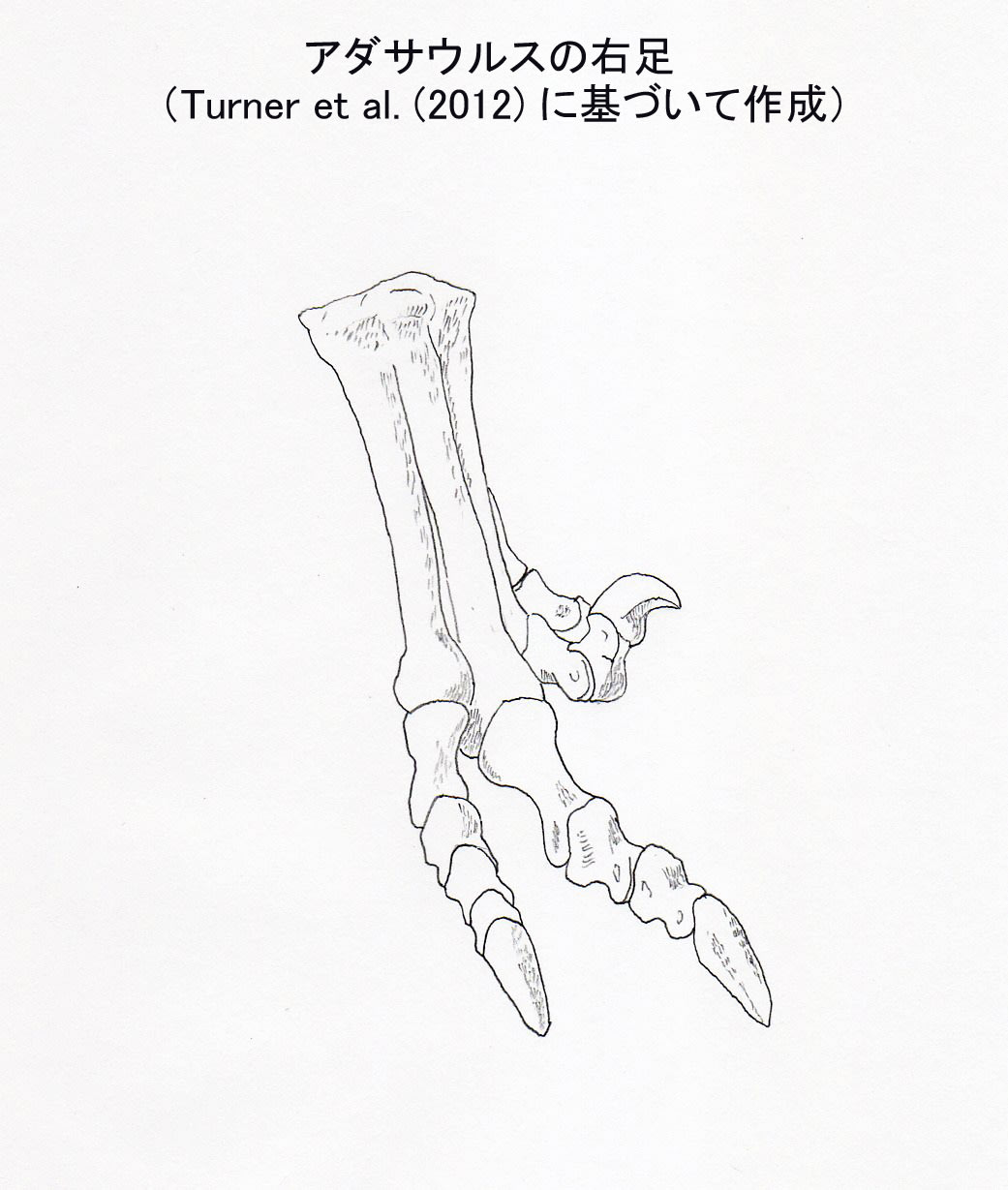
アダサウルスは、白亜紀後期カンパニア期ないしマーストリヒト期(ネメグト層)にモンゴルに生息したドロマエオサウルス類で、Barsbold (1983) によって記載された。ホロタイプIGM 100/20 には頭骨の後半部、脊椎、肩帯、腰帯、後肢の骨が含まれる。パラタイプIGM 100/21 は2個の尾椎と完全な足を含む右後肢からなる。Barsbold (1983)の記載は不十分で、特徴としては後肢の第2指のカギ爪が小さいことしか記述していなかった。またドロマエオサウルス類の中でのアダサウルスの系統的位置ははっきりしていなかった。
アダサウルスの標本は、日本人研究者の久保田克博博士の研究によって再検討された(Kubota and Barsbold, 2006; 2007)。これによりバヤンシレ層から発掘された第3、第4標本(IGM 100/22, IGM 100/23)はアダサウルスではなく、新種のドロマエオサウルス類と考えられた。
Kubota and Barsbold (2006) は、肩甲烏口骨が癒合している点と遠位の足根骨が中足骨と癒合している点で、アダサウルスがヴェロキラプトルと似ていることを見いだした。また他のドロマエオサウルス類と識別できるアダサウルスの特徴として、前頭骨の正中線上に低い稜があること、大きな上角骨孔、腸骨の前寛骨臼突起の前縁に切れ込みがあるnotchedこと、足の第2指のカギ爪が縮小していることを挙げている。ただしこれらの研究はSVPのアブストラクト(学会発表要旨)があるのみで論文にはなっていない。
その後前述のTurner et al. (2012) は、大規模なドロマエオサウルス類の系統研究の中で、アダサウルスのホロタイプ標本を自分で詳細に観察し、多数の新しい形質データをとっている。
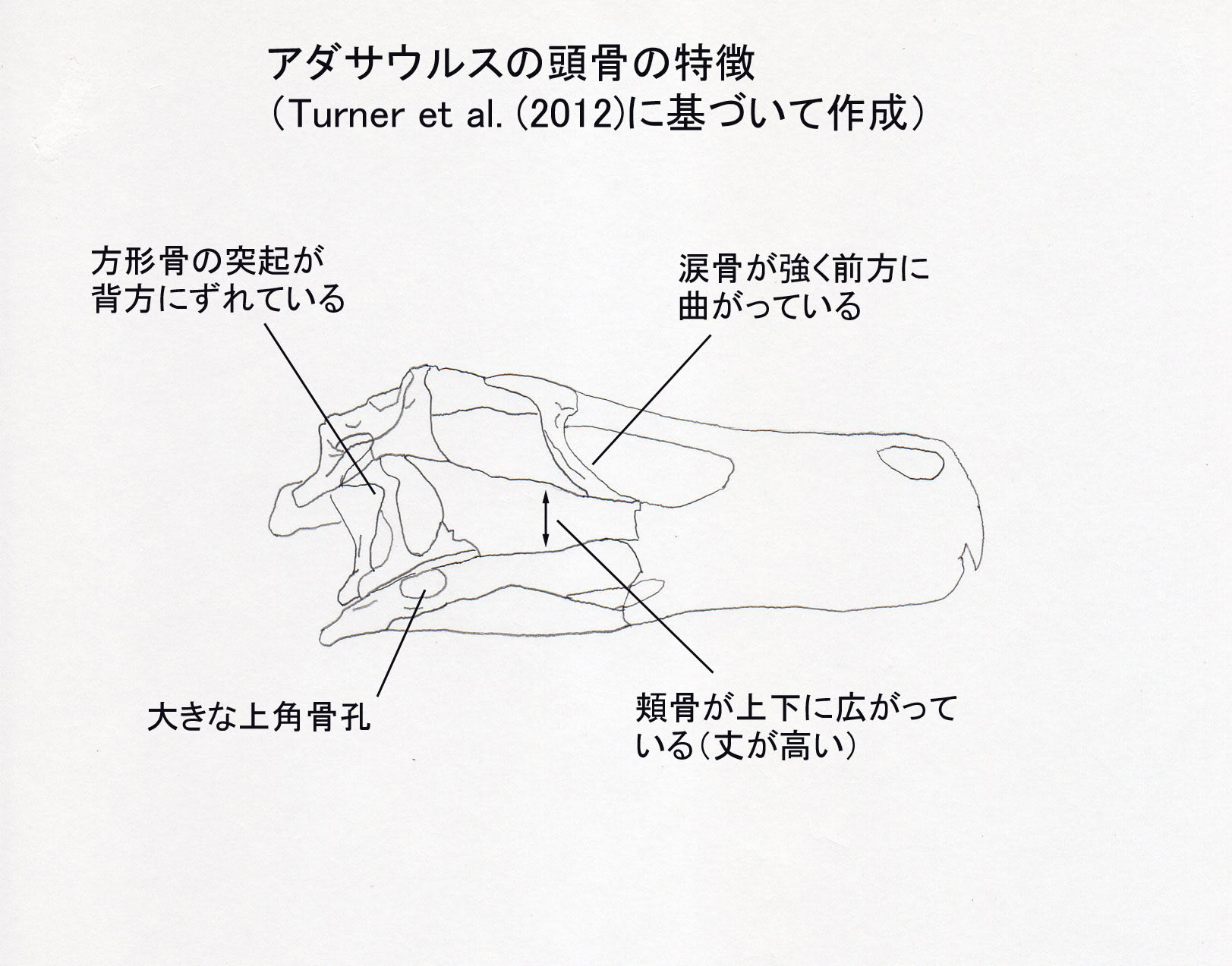
Turner et al. (2012) によると、アダサウルスは以下の形質の組み合わせと固有形質により識別される中型のドロマエオサウルス類である。頬骨の上顎骨突起が上下に拡がっている;涙骨の下方突起(腹側突起)が強く前方に曲がっている;大きな上角骨孔;方形骨の側面にある三角形の突起が背方にずれている;前方の仙椎だけにプレウロシール(側腔)がある;腸骨の前寛骨臼突起の前縁に切れ込みがある;足の第2指のカギ爪が縮小している。
ホロタイプの頭骨は、前半部分、つまり吻部が欠けている。また右側はよく保存されているが、左側は保存がよくない。方形頬骨あたりが欠けているので、方形骨がよく見えている。他のドロマエオサウルス類と同様に、方形骨の側面に大きな三角形の突起がある。この三角形の突起が、他のドロマエオサウルス類では中央あたりにあるが、アダサウルスでは背方にずれているという。涙骨の下方突起が強く前方にカーブしているのは、アダサウルスとアウストロラプトルだけに見られる特徴であるという。Kubota and Barsbold (2006)の挙げた前頭骨の低い稜については、Turner et al. (2012) は、前頭骨の後方部分はプレパレーションが不完全で薄い母岩の層で覆われているため確認できないといっている。
方形骨や下顎の孔は生体復元では見えないので、ヴェロキラプトルやツァーガンと描き分けるのであれば、涙骨のカーブが表現しやすいのではないだろうか。(それも普通には見えないが、色を付けたりして)

大きい画像
アダサウルスの足の第2指のカギ爪は、他の指のカギ爪と同じくらい小さくなっている。「それでもドロマエオかよ」とトロオドンに突っ込まれそうなレベルである。もはや持ち上げなくてもいいような。。。
少し古い恐竜本には、ドロマエオサウルス亜科とヴェロキラプトル亜科の鋸歯の違いが説明してある。Currie (1995) によると、ヴェロキラプトル亜科の特徴は、上顎骨歯と歯骨歯の前縁の鋸歯が後縁の鋸歯よりも顕著に小さいこと、前上顎骨の2番目の歯が3番目と4番目の歯よりも顕著に大きいこと、であった。これはヴェロキラプトルの頭骨を思い浮かべれば比較的わかりやすいと思える。
しかし2001から2003年ごろの系統解析では、ドロマエオサウルス科の内部の系統関係についてほとんどコンセンサスが得られなかった。よってヴェロキラプトル亜科が本当に単系群であるのか、よくわからなかった。その後、Novas and Pol (2005) など、2005から2007年ごろの系統解析によって、ヴェロキラプトル亜科がクレードとして見いだされた。これにはヴェロキラプトル、デイノニクス、ツァーガンが含まれていた。しかしサウロルニトレステスの位置ははっきりせず、ヴェロキラプトル亜科に含まれたり含まれなかったりした。また、このころはアダサウルスはドロマエオサウルス亜科に含まれていた。
Turner et al. (2012) の大規模な系統解析では、ドロマエオサウルス科の中にウネンラギア亜科、ミクロラプトル亜科、ドロマエオサウルス亜科、ヴェロキラプトル亜科に相当するクレードが見いだされた。彼らはアダサウルスの標本を再検討した結果、102箇所もの形質状態を変更した。これによって、アダサウルスは従来のドロマエオサウルス亜科ではなく、ヴェロキラプトル亜科に分類し直された。
Turner et al. (2012) によると、ヴェロキラプトル亜科にはバンビラプトル、ツァーガン、サウロルニトレステス、デイノニクス、ヴェロキラプトル、アダサウルス、バラウルが含まれた。最後の4種は一つのクレードをなしている。
ではアダサウルスのどこがヴェロキラプトル亜科なのかというと、素人にわかりやすいものではないようだ。ヴェロキラプトル亜科は3つの共有派生形質で支持される。基底蝶形骨の後方の開口部が2つの小さな孔に分かれている、dorsal tympanic recessが深く後側方を向いた凹みである、すべての胴椎にプレウロシールがある、ということである。
デイノニクス、ヴェロキラプトル、アダサウルス、バラウルのクレードの共有派生形質は、第 IV 中足骨の骨幹の断面が幅広く扁平であることであるという。
バラウルを除くと、ヴェロキラプトルとアダサウルスは姉妹群をなした。肩甲烏口骨が癒合していること、遠位の足根骨が中足骨と癒合していること、距骨と踵骨が互いに癒合しているが脛骨とは癒合していないこと、の3つの形質で支持されるという。ただし、これらはいずれも骨同士の癒合に関する形質で、成長段階と関連している可能性もある。そこを明らかにするためには、アダサウルスの追加の標本も含めて更なる研究が必要であるという。
参考文献
Kubota, K., and R. Barsbold. (2006) Reexamination of Adasaurus mongoliensis (Dinosauria: Theropoda) from the Upper Cretaceous Nemegt Formation of Mongolia. Journal of Vertebrate Paleontology 26 (suppl. to 3): 88A.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )