獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年7月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | ||||
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| 13 | 14 | 15 | 16 | 17 | 18 | 19 | ||
| 20 | 21 | 22 | 23 | 24 | 25 | 26 | ||
| 27 | 28 | 29 | 30 | 31 | ||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ダスプレトサウルスの系譜(2)

Warshawらの側系統で向上進化説に対する、Scherer and Voiculescu-Holvad (2024) による反論の論拠の一つとして、Brusatte and Carr (2016) などの過去の研究結果と一致しないことが挙げられている。これについてはWarshaw et al. (2024)は、そんなことはなく、ダスプレトサウルスが側系統となる解析結果は過去の研究で繰り返し示されてきたもので、決してとっぴな考えではないと反論している。
もともと発見当初からホルネリ(MOR 590)はダスプレトサウルス・トロススとティラノサウルスの中間的なものとされており、 Horner et al. (1992)の予備的な系統解析でも両者の中間段階となっている、つまりダスプレトサウルスは側系統となっている。本格的なティラノサウルス類の系統解析を行ったHoltz (2001)でも、一部の分岐図ではホルネリはトロススとタルボサウルス+ティラノサウルスの中間段階となった(厳密合意樹では3分岐)。その後Currie (2003)はDinosaur Park Formationのダスプレトサウルスには、トロススと異なる形質を示すものがあることを記載した。Loewen et al. (2013) は初めてトロスス、ホルネリ、Dinosaur Parkのダスプレトサウルスを含めて分岐分析を行った。その結果、ダスプレトサウルスは他のティラノサウルス亜科(テラトフォネウス族とティラノサウルス族)につながる側系統のグレードとなった。Brusatte and Carr (2016) の解析ではテラトフォネウス族がより基盤的でダスプレトサウルスとティラノサウルス族が姉妹群となった。しかしダスプレトサウルスとティラノサウルス族の関係は用いた方法論によって異なり、最尤解析ではダスプレトサウルスが単系統となったが、ベイズ解析ではダスプレトサウルスは側系統となっている。つまりBrusatte and Carr (2016) の解析でさえ、側系統の結果を含んでいるというわけである。
確かに最新のWarshaw et al. (2024)を読むと、単系統のダスプレトサウルスを死守したいのでない限り、ダスプレトサウルスが側系統のグレードになっても別にいいのではないかという気もしてくる。ティラノサウルス族の直接の祖先である動物はカンパニアンのララミディアに生息していたはずである。もしダスプレトサウルスが単系統であるとすると、その中で生じたトロススからホルネリに至る、ティラノサウルス族に近づく変化は真のティラノサウルス族の起源とは結びつかないことになる。それとは別に、ティラノサウルス族の直接の祖先が進化しつつあったことになるが、そういうものは全く見つかってこない。そうではなく、ティラノサウルス族の直接の祖先はすでに見つかっている。それは様々な段階のダスプレトサウルスである、という考え方もありそうな気はするし、ティラノサウルス族の起源を考える上で建設的な気もする。
コメント ( 0 ) | Trackback ( 0 )
ダスプレトサウルスの系譜(1)
これはつい最近も専門家の間で議論が紛糾しているところなので、解説するのは難しいが、研究の流れだけでもまとめておきたい。海外の動画としてはEDGE Science のが短くてよくまとまっているので、それを見れば大体わかる。ダスプレトサウルスには、最近では3種あるいはそれ以上ともされる変異が報告されており、それをどのようにとらえるかについて、ホットな議論が展開されているわけである。
1)ダスプレトサウルス・トロススの発見
ダスプレトサウルスの歴史は、Russell (1970)によるアルバータのOldman Formationからのダスプレトサウルス・トロススの記載から始まった。ホロタイプ標本CMN 8506は完全な頭骨と付属した胴体の骨格からなる。
2)ダスプレトサウルス・ホルネリの記載
Carr et al. (2017) はモンタナ州のTwo Medicine Formationからダスプレトサウルス・ホルネリを記載し、詳細な系統解析の結果、ダスプレトサウルス・トロススとダスプレトサウルス・ホルネリは姉妹群をなすとした。つまりダスプレトサウルスは単系統のクレードであり、その中でトロススからホルネリまではアナジェネシス(向上進化)の例であるとした。向上進化と分岐進化については、すでに2017年の記事に書いてある。
3)ダスプレトサウルス・ウィルソニの記載
Warshaw and Fowler (2022) はモンタナ州のJudith River Formationからダスプレトサウルス・ウィルソニを記載した。ウィルソニは固有派生形質がほとんどなく、トロススとホルネリの中間の形質を示し、生息年代も中間であることから、トロスス、ウィルソニ、ホルネリの順に向上進化を示す系統と考えられた。系統解析の結果、これら3種のダスプレトサウルスはティラノサウルス族(ズケンティラヌス、タルボサウルス、ティラノサウルス)につながる側系統のグレード(段階)と考えられた。つまりダスプレトサウルスは単系統のクレードではなくなってしまうということである。

Copyright 2022 Warshaw and Fowler
4)向上進化と側系統に対する反論
Scherer and Voiculescu-Holvad (2024) は、Warshaw and Fowler (2022)の向上進化説には強い根拠がなく、分岐進化でも説明できるとして、いくつかの新しい標本を加えて独自の系統解析を行った。その結果、4種のダスプレトサウルスが単系統のクレードをなした。またホルネリが最も基盤的で、ウィルソニ、トロススの順に派生的という、逆の系統関係となった。

Copyright 2024 Scherer and Voiculescu-Holvad
5)Warshawらによる再反論―やはり側系統で向上進化である― いまここ
Warshaw et al. (2024) は、Scherer and Voiculescu-Holvad (2024) による反論を受けて、なるべく彼らと同じ形質リストを用い、いくつかの新しい標本を追加し、あらためて系統解析を行った結果、再びダスプレトサウルスは側系統という結果を得た。つまりWarshaw and Fowler (2022) の結果が再確認されることになった。
今回新たに追加されたアルバータのOldman FormationのTMP 2001.36.1、TMP 2003.10.3、モンタナ州Two Medicine FormationのTMDC Daspletosaurus materialは、いずれもトロススとホルネリの中間の形質を示し、ウィルソニに含まれた。
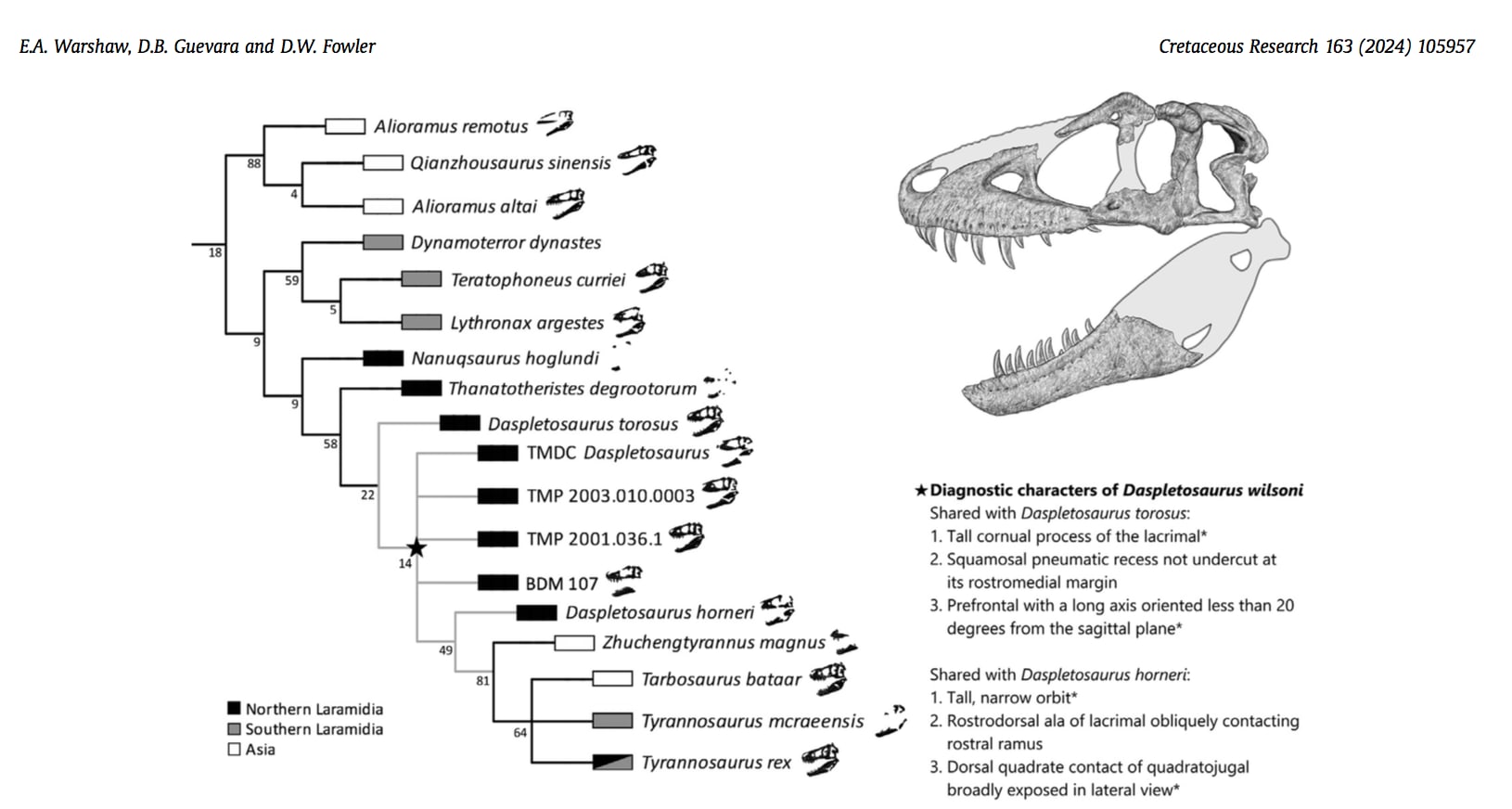
Copyright 2024 Warshaw et al.
今回改訂されたウィルソニの特徴は、まさにトロススとホルネリの中間の形質である。ウィルソニがトロススと共有し、ホルネリにはない形質は例えば涙骨の角状突起が高いことである。これはトロススでは三角形の角状で最も高く、ウィルソニでは中間的でトロススよりは低い角状であり、ホルネリではさらに低くなり尖った角状ではなく丸みを帯びている。また前前頭骨の長軸の角度が矢状面から20°以下であるという。一方、ウィルソニがホルネリと共有し、トロススにはない形質は、上下に高く前後に狭い眼窩である。眼窩の形はトロススではより丸く、ウィルソニでは中間的で、ホルネリではかなり縦長となっている。またウィルソニとホルネリでは、方形頬骨の方形骨との関節部が側面から見えている。
なるほど眼窩の形はかなり異なっている。これを見るとトロススの根本あたりから、テラトフォネウス族が分岐したのではないかとも思えてくる。

(2022の図に 2024 の観点を書き込んだもの)
ダスプレトサウルスが単系統となるか側系統となるかの違いは、Carr et al. (2017)と比較してWarshaw and Fowler (2022)が追加の形質データを用いているからであるという。ダスプレトサウルス・ホルネリの固有派生形質であるとしてCarr et al. (2017)の解析から除外された形質の中に、実は他のティラノサウルス類(特にティラノサウルス族)にもみられる形質が含まれている(Carr et al. もそう記述している)。それらを含めるとホルネリ+ティラノサウルス族というクレードが現れるようである。例えば、上顎骨の最前方の2個の歯間板の形態である。ホルネリとズケンティラヌスは、狭い1番目の歯間板と短縮した2番目の歯間板という形質を共有している。これはウィルソニやトロススとは異なり、またタルボサウルスとティラノサウルスでは1番目、2番目とも幅広くなっている。
Warshaw and Fowler (2022)のときは、ホルネリのデータとしてホロタイプであるMOR 590を用いていた。これに対してCarrらから、MOR 590は亜成体であるなどの批判があったので、今回のWarshaw et al. (2024)ではホルネリのデータとしてMOR 1130(少し時代が新しい、分離した成体の標本)を用いた。その結果、ダスプレトサウルスが側系統であるという結論は変わりなく、むしろ強化されたという。2022のデータでは、上述の歯間板の形質を除くと側系統が崩れてしまい、単系統になってしまう状態だったが、今回の2024ではそのようなことはなく、側系統が保たれる。つまりMOR 1130はいっそうティラノサウルス族に近い形質をもち、ダスプレトサウルスとティラノサウルス族を結びつける要となる標本であるといっている。
参考文献
Elías A. Warshaw, Daniela Barrera Guevara, Denver W. Fowler (2024) Anagenesis and the tyrant pedigree: A response to “Re-analysis of a dataset refutes claims of anagenesis within Tyrannosaurus-line tyrannosaurines (Theropoda, Tyrannosauridae)” Cretaceous Research 163 (2024) 105957
Charlie Roger Scherer, Christian Voiculescu-Holvad (2024) Reanalysis of a dataset refutes claims of anagenesis within Tyrannosaurus-line tyrannosaurines (Theropoda, Tyrannosauridae) Cretaceous Research 155 (2024) 105780
Warshaw EA, Fowler DW. 2022. A transitional species of Daspletosaurus Russell, 1970 from the Judith River Formation of eastern Montana. PeerJ 10:e14461 DOI 10.7717/peerj.14461
Carr, T. D. et al. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Sci. Rep. 7, 44942; doi: 10.1038/srep44942 (2017).
コメント ( 0 ) | Trackback ( 0 )
ラボカニア (想像)

想像とか書かなくても想像図に決まっているが。
眼窩の形をビスタヒ似にして、頭骨はなんとなくテラトフォネウスのようなビスタヒのような感じに。歯の形状から骨を噛むことはなく、小型ないし中型の獲物を狙った(ティラノやタルボだってそうだろうが)。コアフイラケラトプスやトラトロフスと共演させればいいのだろうが、諦めた。
コメント ( 0 ) | Trackback ( 0 )
テラトフォネウス (記念)

テラトフォネウス族樹立記念。このテラトフォネウス族という概念も、もちろん今後の研究で検証されていくものであるが、盛り上がれる時に盛り上がるのが正しい姿勢である。
コメント ( 0 ) | Trackback ( 0 )
ビスタヒエヴェルソル (昇格)

ティラノサウルス亜科テラトフォネウス族に昇格したビスタヒエヴェルソル。「ラボカニアには感謝してる」なにしろダスプレトサウルスよりティラノ・タルボに近いというから大したものである。
原始的な種類ということで推せなかった人も、安心して推して良いだろう。
コメント ( 0 ) | Trackback ( 0 )
ラボカニアの特徴とビスタヒエヴェルソルの目つき


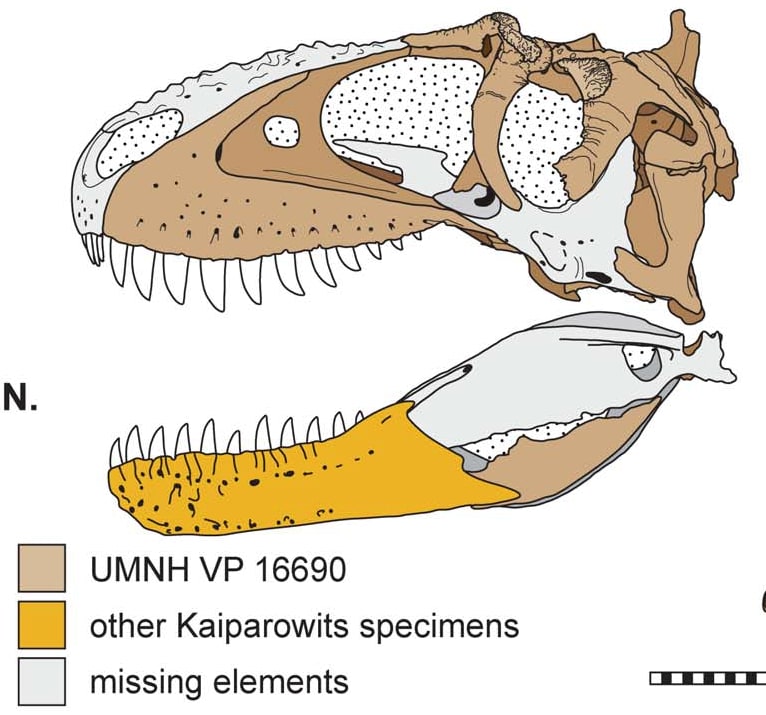
9月に福井で見てきたビスタヒエヴェルソルの頭骨をしみじみ眺めると、なかなかいい顔をしている。改めて見ると、眼窩の形がティラノサウルスやタルボサウルスとはかなり違うことがわかる。またLoewen et al. (2013)を見返すと、同じユタ州でもテラトフォネウスの眼窩の形はリスロナクスと異なることがわかる。
ビスタヒエヴェルソルは、トーマス・カーの系統解析ではティラノサウルス科の外側になり、ティラノサウルス科に入れてもらえなかった。しかしフィリップ・カリー研など他の研究者の間では異論もあった。もっと進化的な種類であり、少なくともティラノサウルス科の中ではないかという意見である。Longrich博士の最近の一連の論文は、ビスタヒエヴェルソルの出世を後押しするものでもある。
ラボカニア・アギロナエLabocania aguillonae は、後期白亜紀カンパニアン後期(Cerro del Pueblo Formation)にメキシコのコアウイラ州に生息したティラノサウルス類で、2024年に記載された。ホロタイプ標本は上顎骨の断片、左右の前頭骨、涙骨の腹側部分、鼻骨の断片、鱗状骨の後端部分など、頭骨、腰帯、後肢にわたるが非常に断片的な骨からなる。論文の全身骨格図を見ればわかるように、大型ティラノサウルス類のシルエットに断片的な骨をあてはめたもので、とても全身像を復元できるようなものではない。
ラボカニアの特徴のうち3つは涙骨に関するもので、これらはいずれもビスタヒエヴェルソルと似て、眼窩が円形に近いことと関連している。1)涙骨の前腹側縁が強く凸型にカーブしている、2)眼窩の前腹側縁に沿って眼窩内に突き出した突起が背側に延びている、3)前眼窩窩が涙骨上を後腹側に広がって、眼窩の前縁の真下で終わっている、である。
左の涙骨の下半分はよく保存されている。涙骨の前縁は強く凸型にカーブしており、ビスタヒエヴェルソルや他のティラノサウルス亜科よりも程度が大きい。この形質はラボカニア・アギロナエの固有派生形質と考えられる。このような涙骨の形から、眼窩の形はビスタヒエヴェルソルのように比較的円形に近いと思われる。一方タルボサウルス、ティラノサウルス、ダスプレトサウルスでは眼窩はもっと縦に長い。
涙骨は腹側で広がっており、そこは後腹側に延びた前眼窩窩で占められている。この部分のよく発達した前眼窩窩はビスタヒエヴェルソル、テラトフォネウス、ダスプレトサウルスにみられる。一方タルボサウルスとティラノサウルスでは前眼窩窩は広がっていない。前眼窩窩は後方に広がり、眼窩の前縁の真下で終わっている。このように前眼窩窩が後腹側に広がることはラボカニア、ビスタヒエヴェルソル、テラトフォネウスの共有派生形質である。ただしこの形質はビスタヒエヴェルソルで最も発達している。ビスタヒエヴェルソルでは、前眼窩窩が眼窩の前縁よりも後方まで広がっている。
涙骨の後縁で、眼窩の前腹側縁に突き出した顕著な結節ventral bossがある。同様の突起はビスタヒエヴェルソルにみられるが、テラトフォネウスや他のティラノサウルス亜科にはない。この突起はラボカニアでは、ビスタヒエヴェルソルよりも背側に広がっているのでラボカニアの固有派生形質と考えられる。
一方、ラボカニアの腸骨と座骨はテラトフォネウスと似ている。腸骨も断片的で寛骨臼の背側縁と恥骨柄pubic peduncleしか保存されていない。恥骨柄は正方形をしている点が特徴的であり、これはテラトフォネウスとよく似ている。また座骨の後背側縁は上方にカーブしており、これはラボカニア・アノマラとテラトフォネウスにみられるが、他のティラノサウルス亜科にもアルバートサウルス亜科にもみられない。
ラボカニア・アギロナエの参照標本として、3個の歯槽が保存された歯骨と多数の分離した歯がある。この歯槽と歯冠の形から、歯冠の側面に溝があり、断面が8の字型の歯をもっていたことがわかった。この特徴はラボカニア・アノマラの歯と一致することから、この参照標本はおそらくラボカニア・アギロナエと考えられる。つまり断面が8の字型の歯は、ラボカニア属に共通した特徴と考えられた。これは獣脚類の中でも珍しい形質であり、多くのティラノサウルス類では断面が直方形か、ティラノサウルスのように太い場合は楕円形である。ちなみにラボカニア・アギロナエのホロタイプ標本は推定6.3 mの亜成体であるが、この参照標本はずっと大きく、ビスタヒエヴェルソルに匹敵する全長8 mと推定された。
系統解析の結果、ラボカニア・アギロナエ、ラボカニア・アノマラ、ビスタヒエヴェルソル、テラトフォネウス、ダイナモテラーは、ティラノサウルス亜科の中でララミディア南部に生息した一つのグループ、テラトフォネウス族Teratophoneiniに含まれた。テラトフォネウス族を識別する特徴には、涙骨、恥骨、座骨の形質が含まれる。また前頭骨と歯の形質はテラトフォネウス族内部のより小さいグループを区別するために用いられる。
ラボカニア・アギロナエの強く円形の眼窩はビスタヒエヴェルソルと共有されるが、ラボカニア・アノマラでは保存されていない。テラトフォネウスでは眼窩は楕円形であり、それほど円形ではない。テラトフォネウスの楕円形からビスタヒエヴェルソルのより円形に近い眼窩への移行は、テラトフォネウス族でより原始的な眼窩の形への逆行が起こったことを示唆している。特徴的な四角形の恥骨柄はテラトフォネウスと共有されるが、ラボカニア・アノマラとビスタヒエヴェルソルでは腸骨が保存されていない。
この研究ではテラトフォネウス族というクレードが存在したことを支持する新しい形質が見いだされた。しかし多くの種類が不完全な標本で知られるのみであることから、さらに追加のより完全な化石の発見と既存のテラトフォネウス族の再研究が必要であると述べている。
参考文献
Rivera-Sylva, H.E.; Longrich, N.R. A New Tyrant Dinosaur from the Late Campanian of Mexico Reveals a Tribe of Southern Tyrannosaurs. Foss. Stud. 2024, 2, 245–272. https://doi.org/10.3390/ fossils2040012
コメント ( 0 ) | Trackback ( 0 )
アジアティラヌス

アジアは広すぎるので、ガンジョウティラヌスとかジャンシティラヌスでよかった。
アジアティラヌスは、後期白亜紀マーストリヒティアン(南雄層Nanxiong Formation)に中国江西省贛州市Ganzhou Cityに生息したティラノサウルス類(ティラノサウルス亜科)で、2024年に記載された。
江西省贛州市の南雄層は恐竜を含む豊富な化石を産出してきたが、獣脚類のほとんどはオヴィラプトル類で、7種も発見されている。それ以外の獣脚類は、吻の細長いアリオラムス族のティラノサウルス類チアンジョウサウルスのみであった。その後、2017年に贛州市の建設工事現場で新たなティラノサウルス類の化石が発見され、浙江自然博物館Zhejiang Museum of Natural Historyでクリーニングされた。吻が短く丈の高いティラノサウルス類としては初めて発見されたものとなった。
アジアティラヌスのホロタイプ標本は、ほとんど完全な頭骨と分離した胴体の部分骨格からなる。尾椎、右の大腿骨、脛骨、腓骨、中足骨、趾骨とより不完全な左後肢である。まあ胴体のほとんどは保存されておらず、頭骨と後肢という感じである。
アジアティラヌスは小型ないし中型のティラノサウルス類で、他のティラノサウルス類と区別される特徴は、前上顎骨の外側面で外鼻孔の近くに2つの小さな深い窪みがある(Fig.6を見る限り左右一対のようである);大きく四角形に近いmaxillary fenestra;鼻骨の後方の突起群が結合して、2つの直列に並んだ正中の稜を形成する(3つか4つの小さい突起がまとまって、ひとかたまりになる。それが2かたまりあって縦に正中線上に並んでいる。おそらくmedium ではなくmedian);頬骨に低い稜状の副次的な角がある;後眼窩骨の下行突起の表面に前背方を向いた線条がある;postorbital barが細くまっすぐで、その前縁と後縁がほぼ平行である、などである。postorbital barは眼窩の後縁の柱状の部分で、後眼窩骨と頬骨からなる。
腓骨の切片の組織像から年齢の推定をしている。13本のLAG(成長停止線)が確認されたことからホロタイプの個体は死亡時に少なくとも13歳であったと思われた。しかし保存が不完全なことから正確なLAGの総数やEFS (external fundamental system)の有無は決定できないという。外側にいくほどLAGの間隔が狭くなっていることや二次的なリモデリングがみられることから、アジアティラヌスのホロタイプは完全に成長した成体ではないが、最も成長が盛んな時期は過ぎていると考えられた。アジアティラヌスの頭骨には、いくつかの成熟した形態学的特徴がみられることから、著者らはこれを成熟に近づいた亜成体と考えている。カンパニアンからマーストリヒティアンの大型ティラノサウルス類では、14歳くらいで指数関数的な成長を示す。それに比べるとアジアティラヌスでは指数関数的な成長の時期が少し早いようである。ホロタイプの大腿骨の長さは、同じような成長段階のゴルゴサウルスなど大型ティラノサウルス類の半分ほどであり、アジアティラヌスは比較的小型のティラノサウルス類と思われる。アジアティラヌスの頭骨の長さは47.5 cmで、全長は3.5-4 mと推定されている。これはチアンジョウサウルスの半分くらいである。
上顎骨の腹側縁は下に凸形にカーブしており、これはティムルレンギアや後期白亜紀の大型ティラノサウルス類と同様で、ほとんどまっすぐなグァンロン、ディロン、シオングァンロン、ススキティランヌスとは異なる。上顎骨の本体はティラノサウルスやタルボサウルスのような大型ティラノサウルス類と同様に短く丈が高く、アリオラムス・レモトゥス、アリオラムス・アルタイ、チアンジョウサウルスの長く丈の低い上顎骨とは異なる。
上顎骨の外表面には、ダスプレトサウルス、タルボサウルス、ティラノサウルス、ズケンティラヌスにみられるような背腹方向の溝や稜はみられない。上顎骨と前上顎骨の関節面は多くのティラノサウルス類と同様に背腹方向を向いており、関節面が強く後背方に傾いたアリオラムス族とは異なる。いくつかの神経血管孔が歯列の背側に並んでいる。
Maxillary fenestra は大きく四角形に近い形であり、これはタルボサウルスの成体と似ているが、タルボサウルスの幼体では楕円形である。他のティラノサウルス亜科と異なり、maxillary fenestra の前縁は前眼窩窩の前縁とは接していない。それでもmaxillary fenestraは前眼窩窩の中で比較的前方に位置している。一方、アリオラムス・アルタイ、アパラチオサウルス、アルパートサウルス、ビスタヒエヴェルソル、ゴルゴサウルス、ティラノサウルス亜科の幼体では、maxillary fenestraがより中央に位置している。上顎骨の歯列は完全には保存されていない。
涙骨の角状突起は、ゴルゴサウルスのような尖った角状ではなく、全体に膨張したinflatedゆるやかな山形である。これはティラノサウルスとタルボサウルスにみられる状態である。この膨張inflationはラプトレックスにはみられず、またタルボサウルスの幼体ではあまり発達していない。角状突起が尖った角をなすのは、ユーティラヌス、アルバートサウルス亜科、ダスプレトサウルス、アリオラムス・アルタイ、チアンジョウサウルスであるといっている。
論文の全身骨格図をみると、後眼窩骨の眼窩下突起suborbital processがあるように見える。しかしこの描き方が不正確で、後眼窩骨の記述を読むと眼窩下突起はないとはっきり書いてある。皮膚をつければ隠れるとはいえ、注意が必要である。
アジアティラヌスでは後眼窩骨の下行突起はアリオラムス・アルタイやナノティラヌスと似て舌形で細長い。また下行突起はまっすぐで、前方に広がった眼窩下突起はない。多くの大型ティラノサウルス類(ティラノサウルス、タルボサウルス、アルバートサウルス、ゴルゴサウルス)では下行突起が広がって眼窩内に突き出した眼窩下突起をもつ。それに対して、アジアティラヌスの下行突起の前縁はほとんどまっすぐである。後眼窩骨と頬骨からなるpostorbital barは細長くまっすぐで、前縁と後縁が平行になっている。他の眼窩下突起をもたないティラノサウルス類(ダスプレトサウルスやナノティラヌス)では、下行突起の前縁はむしろ凹形にカーブしている。
系統解析の結果、アジアティラヌスはティラノサウルス科のティラノサウルス亜科に含まれた。アジアティラヌスはティラノサウルス亜科の中では、アリオラムス族やテラトフォネウスよりは派生的で、ダスプレトサウルスよりは基盤的な位置にきた。これはナヌークサウルスと近い段階である。ナヌークサウルスも推定頭骨長60-70 cmと小型ないし中型であるが、最近の知見によるとナヌークサウルスはもっと大きかった可能性があるという。そうするとアジアティラヌスは唯一、確実に小型のティラノサウルス亜科ということになる。
アジアではモンゴルのタルボサウルスとアリオラムス、中国南部のチアンジョウサウルスとアジアティラヌスというように、吻の丈が高いティラノサウルス類と吻の細長いティラノサウルス類が共存していたことがわかってきた。しかし中国南部では吻の細長いチアンジョウサウルスの方が大型で、体の大きさはモンゴルとは逆転している。吻の形状が異なることから、両者は異なるニッチを占めて棲み分けていただろうとしている。大型だが吻が細長いものと、顎が頑丈だが小型のものでは、どちらも小型の獲物を狙ったような気がするがどうなのだろうか。
参考文献
Wenjie Zheng, Xingsheng Jin , Junfang Xie & Tianming Du (2024) The first deep‑snouted tyrannosaur from Upper Cretaceous Ganzhou City of southeastern China.
Scientific Reports (2024) 14:16276 | https://doi.org/10.1038/s41598-024-66278-5
コメント ( 0 ) | Trackback ( 0 )

ティラノサウルス・マクラエーンシスとシエラケラトプス
ティタノサウルス類が来るのはかなり後なので、同時には描きにくいかと。
コメント ( 0 ) | Trackback ( 0 )
ティラノサウルス・マクラエーンシス

Copyright 2024 Dalman et al.
ついこないだもグレゴリー・ポールの3種説を、トーマス・カーら専門家が全力で否定したばかりである。ティラノサウルスが何種あるのかという問題については、今後もなかなか議論が尽きないのだろう。しかしそれは別にしてもティラノサウルスの起源がどこなのかという問題について、大きく貢献する研究である。
Scientific Reportsの論文はコンパクトにまとまっているし、海外の動画も3つくらい上がっている。よって私が紹介する立場にないので詳しくは書かない。
ティラノサウルス・マクラエーンシスTyrannosaurus mcraeensis は、カンパニアン末期からマーストリヒティアン初期(Hall Lake Formation)に、ニューメキシコ州シエラ郡Sierra Countyに生息したティラノサウルス属の新種で、2024年に記載された。ティラノサウルス・レックスよりも600-700万年も古い生息年代ながら、ティラノサウルス・レックスと同等の大きさ12 mであったと推定され、ティラノサウルスの大型化がララミディア南部で起きたことを示唆している。
ティラノサウルス・マクラエーンシスのホロタイプ標本NMMNH P-3698は部分的な頭骨で、右の後眼窩骨と鱗状骨、左の口蓋骨、上顎骨の断片、下顎(左の歯骨、右の夹板骨(板状骨)、前関節骨、角骨、関節骨)、分離した歯、血道弓からなる。
ティラノサウルス・マクラエーンシスの特徴は、後眼窩骨の角状突起が低く後方に位置するなど13くらいあるが、ティラノサウルス・レックスとの違いを示した図をみるのがわかりやすい。
後眼窩骨にはティラノサウルス亜科に典型的な、背方に突き出した大きな角状突起cornual process, cornual bossがある。ティラノサウルス・レックスでは角状突起が前方に強く膨らんで、その頂点が前方、つまり眼窩の上にある。それに対してティラノサウルス・マクラエーンシスでは角状突起が前方に膨らんでおらず、頂点がより後方にある。
また後眼窩骨の前頭骨・前前頭骨との関節面(のある突起)は、ティラノサウルス・マクラエーンシスでは前方を向いているが、ティラノサウルス・レックスでは前腹方を向いている。
最もわかりやすいのは歯骨の下側のラインである。ティラノサウルス・レックスでは下顎のつけねがぐぐっと膨らんでいるので、歯骨の後端部は太く、後腹側縁が下がっている。それに対してティラノサウルス・マクラエーンシスでは歯骨の後端部は丈が低く、後腹側縁のラインはむしろ上がっている。つまり下顎の下縁のラインがほとんどまっすぐになっている。これはティラノサウルス亜科の中でもユニークな特徴であるが、タルボサウルスやズケンティラヌスにより近いという。この歯骨の大きさから、ティラノサウルス・マクラエーンシスはスコッティのような最大級のティラノサウルスよりは小さいものの、ティラノサウルス・レックスと同等の大きさと推定されている。歯骨の歯槽の数は13で、ティラノサウルス・レックスと同様である。ズケンティラヌスでは15,タルボサウルスでは14-15,ダスプレトサウルス・ホルネリとダスプレトサウルス・トロススでは17である。歯骨の前端の下顎結合symphysisの部分はティラノサウルス・レックスと同様に丈が高い。この部分は歯骨の腹側縁から急に立ち上がっているので角ばったおとがいをなしているのもレックスと同様である。
系統解析の結果、ティラノサウルス・マクラエーンシスはティラノサウルス・レックスと姉妹群となり、これら2種はアジアのティラノサウルス亜科であるタルボサウルスとズケンティラヌスのクレードと姉妹群となった。
ティラノサウルス・マクラエーンシスをティラノサウルス・レックスと区別する特徴は比較的微妙なものであるが、後者では多数の個体が知られているので、その個体変異の範囲と比べてどうなのか、が検討できる。その結果、今回示されたティラノサウルス・マクラエーンシスの特徴は、ティラノサウルス・レックスのどの標本とも異なるものであるとしている。またマクラエーンシスの標本はレックスの成体と同等の大きさなので、成長段階による違いとは考えられない。また著者らは、マクラエーンシスとレックスの違いはそれぞれの骨について1つ以上あることも強調している。
ティラノサウルス・レックスはマーストリヒティアン後期に突然現れているため、その祖先についてはわかっていなかった。最も近縁な種類がアジアのタルボサウルスとズケンティラヌスであることから、祖先がアジアに移動し、その後北米に戻ったという説と、北米に留まって進化したという説があった。今回の系統解析の結果からみると、ティラノサウルスの祖先はララミディア南部に出現し、一つの系統はアジアに渡ってそこでタルボサウルスとズケンティラヌスを生み出した。もう一つの系統は北米で地域固有の進化を遂げ、ティラノサウルスとしてララミディア北部に進出した、というシナリオが考えられる。
ニューメキシコ州のHall Lake Formation がかつてマーストリヒティアン後期と考えられていた理由の一つは、ティラノサウルス・レックスとトロサウルスの存在であった。以前トロサウルスと思われていた角竜は、現在、新種のカスモサウルス類シエラケラトプスとされている。シエラケラトプスは、カンパニアン末期のコアフイラケラトプスなどと近縁とされている。
Hall Lake Formationの恐竜相には、大型のティラノサウルス・マクラエーンシス、大型のカスモサウルス類シエラケラトプス、アラモサウルスと比較されるティタノサウルス類、大型のハドロサウルス類が含まれていた。これらは同時代の北部ララミディア(カナダ)の恐竜相とは大きく異なっている。当時カナダには、アルバートサウルス亜科アルバートサウルス、セントロサウルス亜科パキリノサウルス、ランベオサウルス類ヒパクロサウルス、ハドロサウルス類エドモントサウルスなどがおり、竜脚類はいなかった。
ララミディア南部では大型の植物食恐竜が繁栄しており、それに適応して大型のティラノサウルスが進化し、やがてララミディア北部にも広がった可能性が考えられた。
例によってこのティラノサウルスの新種が今後も認められるかどうかはまだわからない。しかし仮に別種ではなくても、このような古い時代からティラノサウルスが存在していたということは、マーストリヒティアン末期の恐竜というティラノサウルスの概念に大きな影響があるに違いない。
参考文献
Sebastian G. Dalman, Mark A. Loewen, R. Alexander Pyron, Steven E. Jasinski, D. Edward Malinzak, Spencer G. Lucas, Anthony R. Fiorillo, Philip J. Currie & Nicholas R. Longrich (2024) A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism. Scientific Reports (2024) 14:22124
コメント ( 1 ) | Trackback ( 0 )
| « 前ページ |