獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年9月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
リンヘヴェナトル

(未完成)
リンヘヴェナトルは、白亜紀後期カンパニアン(Wulansuhai Formation)に中国内モンゴル自治区のBayan Mandahuに生息した前肢の短いトロオドン類で、2011年に記載された。
ホロタイプLHV0021は一部が関節した部分骨格で、頭蓋、下顎、6個の前方と中央の胴椎、右の肩甲骨と右の上腕骨、不完全な左右の座骨、左大腿骨、ほとんど完全な左足、その他の断片的な骨からなる。化石の保存状態は良くなく、ひどく風化していて、頭骨の骨の表面には浸食による多数の溝や稜があり、胴体の骨の関節端も浸食されている。それでも白亜紀後期の進化したトロオドン類の骨格としては、よく保存されている方だという。
神経弓と椎体の癒合から恐らく成体と考えられ、大腿骨の長さが240 mm、推定体重23 kgと、比較的大きなトロオドン類であるという。頭骨の長さは220 mmと推定され、これはサウロルニトイデスのホロタイプと同じくらいであるという。
全身の部分骨格なのでそれなりに情報が得られたと思われるが、頭骨は完全につぶれていて上顎や下顎の形態情報は得られない。それでも保存のよい足がアルクトメタターサルであることと、分離した歯があったことで、専門家にとってはやりやすい方かもしれない。
リンヘヴェナトルは、後肢の第II指の特殊化した形態から、明らかにデイノニコサウリアに属する。またリンヘヴェナトルはいくつかの共有派生形質に基づいて、トロオドン類に分類される。鼻骨と涙骨が吻の上でlateral shelfを形成する、涙骨の下行突起の上にlateral flangeがある、涙骨の前方突起が長くのびて前眼窩窓全体の背側縁をなす、中足骨が非対称で、第II中足骨が短く細く、第IV中足骨が太い、である。
トロオドン類の中で、リンヘヴェナトルは完全にアルクトメタターサルな中足骨をもつことと、典型的なトロオドン類の歯(カーブした低い歯冠、基部がくびれている、鉤状の大きな鋸歯)をもつ点で、シノヴェナトルやメイのような基盤的な種類よりも派生的である。
系統解析の結果、リンヘヴェナトル、トロオドン、ザナバザル、サウロルニトイデスが派生的なトロオドン類のクレードを形成した。リンヘヴェナトルがトロオドンと似ている点として、三角筋稜の縁にある稜や、比較的中足骨ががっしりしていることがあげられるという。
トロオドン類の中でも、リンヘヴェナトルとトロオドンでは後肢の第II指のカギ爪がドロマエオサウルス類とよく似て、大きく発達している。つまり指骨II-1にはわずかにヒールがあり、指骨II-2は高度に短縮して大きなヒールがあり、末節骨は他の指よりも顕著に大きくなっている。これらの形態は基盤的なトロオドン類よりもドロマエオサウルス類に近づいてきている。また恐らくそれと関連して、リンヘヴェナトルとトロオドンの中足骨は基盤的なトロオドン類よりも短く太くなっている。これらの特徴からドロマエオサウルス類と派生的なトロオドン類は、捕食のための高度に特殊化したカギ爪を独立して進化させたと思われる。
これまで知られているトロオドン類は、近縁のドロマエオサウルス類やアヴィアラエよりは短い前肢をもっているが、トロオドン類の前肢は獣脚類一般の中では特に短いわけではない(上腕骨/大腿骨の比が0.52 から 0.65)。リンヘヴェナトルの上腕骨/大腿骨の比は0.4で、獣脚類の中でもかなり短い。前肢の短いドロマエオサウルス類アウストロラプトルでは0.46、コンプソグナトゥスでは0.44、ティラノサウルスでは0.29、アウカサウルスでは0.35である。
派生的なトロオドン類では上腕骨と大腿骨の両方が保存された標本はほとんどないので、他の派生的なトロオドン類の前肢がリンヘヴェナトルのように短かったかどうかはわからない。ただしトロオドンの断片的な前腕と手の化石には、それを示唆するものがあるという。もしかすると派生的なトロオドン類には、前肢が短くなる進化傾向があったのかもしれないが、それを検証するためにはもっと多くのトロオドン類の四肢の標本が必要である。
リンヘヴェナトルの前肢が短いことは、捕食にあまり前肢を用いなかったことを示唆する。しかしリンヘヴェナトルの上腕骨は、短いががっしりしていて、三角筋稜などの筋付着部が大きく発達していることから、何らかの機能を果たしていたことは確かであるという。著者らは捕食のほか、穴を掘る、よじのぼるなどを挙げているが、リンヘヴェナトルが前肢をどのように使ったかはわからない。ちなみにアウストロラプトルでは上腕骨の大きさのわりに三角筋稜の発達は悪いという。
参考文献
Xu X, Tan Q, Sullivan C, Han F, Xiao D (2011) A Short-Armed Troodontid Dinosaur from the Upper Cretaceous of Inner Mongolia and Its Implications for Troodontid Evolution. PLoS ONE 6(9): e22916. doi:10.1371/journal.pone.0022916
コメント ( 0 ) | Trackback ( 0 )
シノルニトイデス

サウロルニトイデス、シノルニトイデスと聞くと、JALの「うどんですかい」「そばですかい」を思い出す。しかし「ちゃんぽんですかい」も出ていたとは知らなかった。オルニトミムス類は「なんとかミムス」、アルヴァレスサウルス類は「なんとかニクス」のように、トロオドン類の名前も「なんとかオイデス」で統一されていれば覚えやすかった。残念ながらトロオドン類はバラバラで、最近はヴェナトルが増殖している。ヴェナトルでは何の仲間かわからない。
シノルニトイデスは、白亜紀前期(Ejinhoroqi Formation)に中国内モンゴルのオルドス盆地に生息した小型のトロオドン類で、1993年に記載された。全長1mほどの小型ながら、発見当時は最も完全なトロオドン類の全身骨格として注目されたはずである。いま見てもすごい化石には違いない。シノルニトイデスの発見により、方形骨が脳函に接している、胸骨が骨化していないなど、それまで知られていなかったトロオドン類の特徴が明らかになった。また全身の骨格が復元されたことで、後肢が非常に長く、その割に前肢は短いというトロオドン類のプロポーションが示された。これを参考にトロオドン類の復元画を描いた人も多いだろう。
メイの第二標本の論文に書いてあるが、実はシノルニトイデスは、メイより早く発見された「元祖・眠り竜」である。メイと同じ休息姿勢をとっている。地面に腹をつけて座り、尾を体に巻き付けるように曲げ、折りたたんだ前肢の上に頭がくるように首を曲げている。手の指の角度までメイと同様であるという。残念ながら体の背側にあった頭骨の右半分、頸椎の一部、ほとんどの胴椎などは侵食されて失われている。
多くの恐竜が死亡時には背中を反らせた姿勢を残しているが、これは窒息、毒物、高熱などに対する生体反応で、「断末魔のあがき」の姿勢であるという意見があるという。メイやシノルニトイデスがそうならず、安らかに眠っていることについて、あがきの生体反応を起こす間もなく堆積物に埋もれた、有毒な火山ガスなどによって生体反応が阻害された、などの可能性が考えられている。遼寧省では火山灰、内モンゴルでは砂嵐などによって一瞬で埋められたのだろうか。
今回はトロオドン類の顔つきを比較するのが趣旨であったが、見えないものは仕方がない。シノルニトイデスの頭骨は、腹側からみて前肢と尾椎の陰に隠れていて、うまく観察できない。シノルニトイデスの記載論文はRussel and Dong (1993)、 Currie and Dong (2001)の2つあるが、どちらも頭骨について結構細かいことを記述しているわりには、頭骨の拡大写真というものがない。解像度のあまりよくない全身の白黒写真があるのみである。少なくとも上顎や下顎周辺は保存されているはずで、上顎骨の孔や歯の数などの形態を論じているのに写真も図もなく、なんとももどかしい。Currie and Dong (2001)の中に歯と歯骨の前端の図はある。また侵食によって分離していた前頭骨の図はある。眼窩から後方は部分的に保存されているが、はっきりした形態はわからないらしい。眼窩内に強膜輪がある、鱗状骨に孔がある、方形骨が脳函に接しているなどの記述は図示されていない。
1993年当時にはトロオドン類は、トロオドン、サウロルニトイデス(2種)、ボロゴヴィア、未命名のトロオドン類(Barsbold et al. 1987)のわずか4属しか知られていなかった。2001年でもそれほど増えておらず、その後になってシノヴェナトル、メイ、シヌソナスス、ウルバコドンなど重要な種類が続々と発見されている。そのため1993年の記載論文では、シノルニトイデスの特徴として、トロオドンやサウロルニトイデスなどとの細かい違いがあげられている。
他のトロオドン類と区別されるシノルニトイデスの特徴は、
1)サウロルニトイデスよりも頭骨が小さく、近位の尾椎や足の趾骨との相対値で比較すると、サウロルニトイデスの0.7 倍である。シノルニトイデスの頭骨はサウロルニトイデスの半分しかないが、足の骨は平均してサウロルニトイデスの80%以上に達する。つまりより大型のトロオドン類であるトロオドンやサウロルニトイデスでは、もっと頭骨が大きいということである。
2)上顎骨と鼻骨の縫合線が、上顎骨の腹側縁に対して20°傾いている。(これは他のトロオドン類ではどうなのか説明はない。)
3)側頭部が眼窩よりも狭い。これはサウロルニトイデスと同様で、トロオドンとは異なる。
4)前頭骨の眼窩の縁が、正中側の縁に対して20°傾いている。これはトロオドンやサウロルニトイデスでは40°である。つまりシノルニトイデスの方がより側方を向いている。
5)脛骨の長さは大腿骨の1.4倍である。
6)第III中足骨の遠位の関節面の後腹側に舌状tongue-likeの突出部があまり発達していない。これは白亜紀後期のトロオドン類にはみられる。
7)ボロゴヴィアにある足の特殊化はみられない。
(現在では、全身骨格が保存されたものを含め、はるかに多くのトロオドン類が蓄積しているので、保存のよい四肢や後半身にシノルニトイデスの特徴があるのかもしれない。最近の論文や総説の中に記述があるかもしれないが、すべて調べてはいない。)
シノルニトイデスは白亜紀前期のトロオドン類なので、一部の形質については白亜紀後期のトロオドン類よりも原始的であるが、すでに多くのトロオドン類の特徴をもっている。前上顎骨はあるが、internarial barについては情報がない。外鼻孔の腹側縁には上顎骨が参加している。涙骨の前方突起は保存されていないが、下行突起の上のflange (shelf) はあると考えられている。歯骨の側面の溝の中に神経血管孔が並んでいる。上顎の歯列は前方で密集しており、後方で間隔があいている。10番目以後の尾椎では、神経棘がなくなっている。
歯は比較的小さく数が多く、歯冠と歯根の間が顕著にくびれている。前上顎骨歯が4、上顎骨歯が18ある。歯骨歯は観察できるものが12あるという。前上顎骨歯は、前縁と後縁の両方に鋸歯がある。上顎骨歯と歯骨歯は、後縁のみに鋸歯がある。これはサウロルニトイデスと同様で、トロオドンとは異なる。トロオドンやサウロルニトイデスの歯との重要な違いは、シノルニトイデスの歯では鋸歯が比較的小さいことである。歯冠の大きさを横軸に、鋸歯のサイズ(denticular basal diameter)を縦軸にとって対数グラフを描くと、他のテタヌラ類の歯の回帰直線よりも上に、トロオドン類の回帰直線がくる。その中でシノルニトイデスの歯は最も下にプロットされる。ただし、一般の獣脚類よりは大きいという。つまりシノルニトイデスの鋸歯は、一般の獣脚類よりは大きいが、進化したトロオドン類よりは小さい。(この時は発見されていないがシノヴェナトルの鋸歯は小さいはずである。)
続くかも
参考文献
Russell DA, Dong ZM (1993) A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People’s Republic of China. Can J Earth Sci 30: 2163-2173.
Currie PJ, Dong Z (2001) New information on Cretaceous troodontids (Dinosauria, Theropoda) from the People’s Republic of China. Can J Earth Sci 38: 1753-1766.
コメント ( 0 ) | Trackback ( 0 )
ウハートルゴドの ”サウロルニトイデス”
最近のトロオドン類の記事を自分で読み返している時に、一見矛盾があることに気づいたので、調べて補足説明します。
メイの記事中で、尾椎の溝は「サウロルニトイデス、‥‥にもみられる」とあるのは、確かにGao et al. (2012) に書いてあるが、このサウロルニトイデスは別物です。サウロルニトイデスのホロタイプ以外に、歴史的にサウロルニトイデスと呼ばれた標本はいくつかあり、その一つがNorell and Hwang (2004) が記載したモンゴルのウハートルゴドの標本です。
これは部分的な上顎骨、方形骨、頚椎、尾椎、中足骨などからなる断片的な化石で、上顎骨の歯列や表面の溝のパターンなどがサウロルニトイデスに似ていることなどから、暫定的にサウロルニトイデス・モンゴリエンシスとされたもので、あくまでcf. Saurornithoides mongoliensis です。後のNorell et al. (2009) はホロタイプ以外は認めていないので、正式にはサウロルニトイデスとはいえないものです。よってメイの記事中では 括弧付きの”サウロルニトイデス”に訂正します。
この ”サウロルニトイデス”には、3つの後方の尾椎が含まれていて、神経棘がなく背側の溝が走っているとある。残念ながら写真は載っていない。典型的なトロオドン類の形態ということでしょう。
この ”サウロルニトイデス”は非常に断片的ではあるが、ホロタイプより少し大きい。これが持つトロオドン類の特徴は、アルクトメタターサルな足(分離した中足骨から)、上顎骨の歯列が密集していること、外鼻孔の縁に上顎骨が面していること、などである。
上顎骨には5本の歯根が保存されていて、その5本は密集していて境界が不完全、つまりつながった溝に収まっている。最後の5番目の歯の後方には薄いinterdental boneの壁があるので、6番目以降は仕切られた歯槽に収まっていると考えられた。これも典型的なトロオドン類の特徴である。
また栄養孔や置換孔という孔を通して歯根が観察できるが、成熟した歯の真下、歯髄腔の位置に次の成長中の歯が収まっている。これは、ワニ類やアヴィアラエと同様の垂直交換vertical replacement であるといっている。ほとんどの獣脚類では成熟した歯の内側(舌側)に次の歯が萌出する形になるのに対し、トロオドン類では鳥類と同様の垂直交換様式をとることがわかった、としている。
歴史的にサウロルニトイデスと呼ばれた標本のもう一つは、Currie and Peng (1993) のサウロルニトイデスの幼体とされた小さい足で、これは現在、フィロヴェナトルとなっている。
参考文献
Norell MA, Hwang S (2004) A troodontid dinosaur from Ukhaa Tolgod (Late Cretaceous Mongolia). Am Mus Novit 3446: 1–9.
コメント ( 0 ) | Trackback ( 0 )
由緒正しいサウロルニトイデス

サウロルニトイデス・モンゴリエンシスは、白亜紀後期カンパニアン(Djadokhta Formation)にモンゴルのバヤン・ザクBayan Zagに生息した中型のトロオドン類で、Osborn (1924) によって記載された。その当時、トロオドンの歯は発見されていたが獣脚類と認識されていなかったので、サウロルニトイデスは最も由緒正しいトロオドン類である。実際にトロオドン科は一時期、サウロルニトイデス科と呼ばれたことがある。
その後ブギン・ツァフのネメグト層から保存のよい頭骨を含む部分骨格が発見され、Barsbold (1974) によりサウロルニトイデス・ジュニアSaurornithoides juniorと命名された。頭骨の特徴がサウロルニトイデス・モンゴリエンシスと似ていることと、モンゴルの上部白亜層から産することから同属とされたものである。しかしさらにその後、Norell et al. (2009)が再検討した結果、これら2種はトロオドン類の中で近縁ではあるが、同属とするほどではないと考えられ、サウロルニトイデス・ジュニアは新属ザナバザル・ジュニアとなった。
サウロルニトイデスのホロタイプは、ほとんど完全だが侵食された頭骨、一連の胴椎、仙椎、尾椎、部分的な腰帯と後肢の骨からなる。
Norell et al. (2009) はサウロルニトイデスとザナバザルを比較・再検討している論文なので、サウロルニトイデスの特徴は、最も近縁なザナバザルとの違いで表現されている。サウロルニトイデスに固有の形質があまり表現されていないようにみえるが、これでいいのだろうか。
サウロルニトイデスとザナバザルの違い
1)大きさ:最も明らかな違いは全体の大きさで、サウロルニトイデスの方が小さい。頭骨の長さはサウロルニトイデスが189 mm に対してザナバザルが272 mm である。これは1個体同士の比較であるが、どちらも成体である。
2)歯の数:サウロルニトイデスの方が歯の数が少ない。サウロルニトイデスでは上顎骨歯19 、歯骨歯31-33 であるが、ザナバザルでは上顎骨歯19-20 、歯骨歯35 である。
3)頰骨の形:頰骨の眼窩の下の部分がザナバザルではカーブしているが、サウロルニトイデスではまっすぐである。
4)脳函の窪み:サウロルニトイデスでは、脳函の前耳骨の側面で三叉神経孔の背側に、含気性の窪みがある。この窪みはザナバザルにもトロオドンにもない。ただしこの窪みはビロノサウルスやシノヴェナトルにもみられるので、派生形質ではなく原始形質と思われる。
5)サウロルニトイデスの上顎骨歯は後方へいくにつれて大きくなるが、その程度はザナバザルの場合より小さい。
6)サウロルニトイデスの上顎骨歯には交代ギャップreplacement gap がないが、ザナバザルにはある。
著者らは5)6)の点について、サウロルニトイデスはメイやシノヴェナトルと似ており、ザナバザルはビロノサウルスと似ていると言っている。
サウロルニトイデスは派生的なトロオドン類なので、多くのトロオドン類の特徴を一通り備えているようである。下顎はひどく侵食されているので、歯骨の側面の溝は後方 1/3 くらいの部分のみ明確に観察される。また下顎は上顎の内側にはまり込んでいるので、下顎の歯列は内側からのみ観察できる。歯列は上顎も下顎も同じ傾向を示すのであるが、歯骨の先端の歯は保存されていないようだ。上顎骨歯と歯骨歯は典型的なトロオドン類の歯であり、後縁のみに大きな鋸歯がある。
後肢の骨のうち、足は部分的にしか保存されていないが、第II中足骨が細く第IV中足骨が太い、第III中足骨は近位で細くなっているなど、非対称でアルクトメタターサルと考えられる特徴を示す。第II趾のカギ爪はほどほどに発達している。後方の尾椎は保存されていないので、背側の溝などの情報はない。
サウロルニトイデスの顔は細長いが、意外としっかりしている印象がある。もう少し保存が良ければ、ザナバザルに負けない標本になっていたような気がする。
参考文献
Norell MA, Makovicky PJ, Beaver GS, Balanoff AM, Clark JM, et al. (2009) A Review of the Mongolian Cretaceous dinosaur Saurornithoides (Troodontidae: Theropoda). Am Mus Novit 3654: 1-63.
コメント ( 0 ) | Trackback ( 0 )
鋸歯のないトロオドン類ビロノサウルス
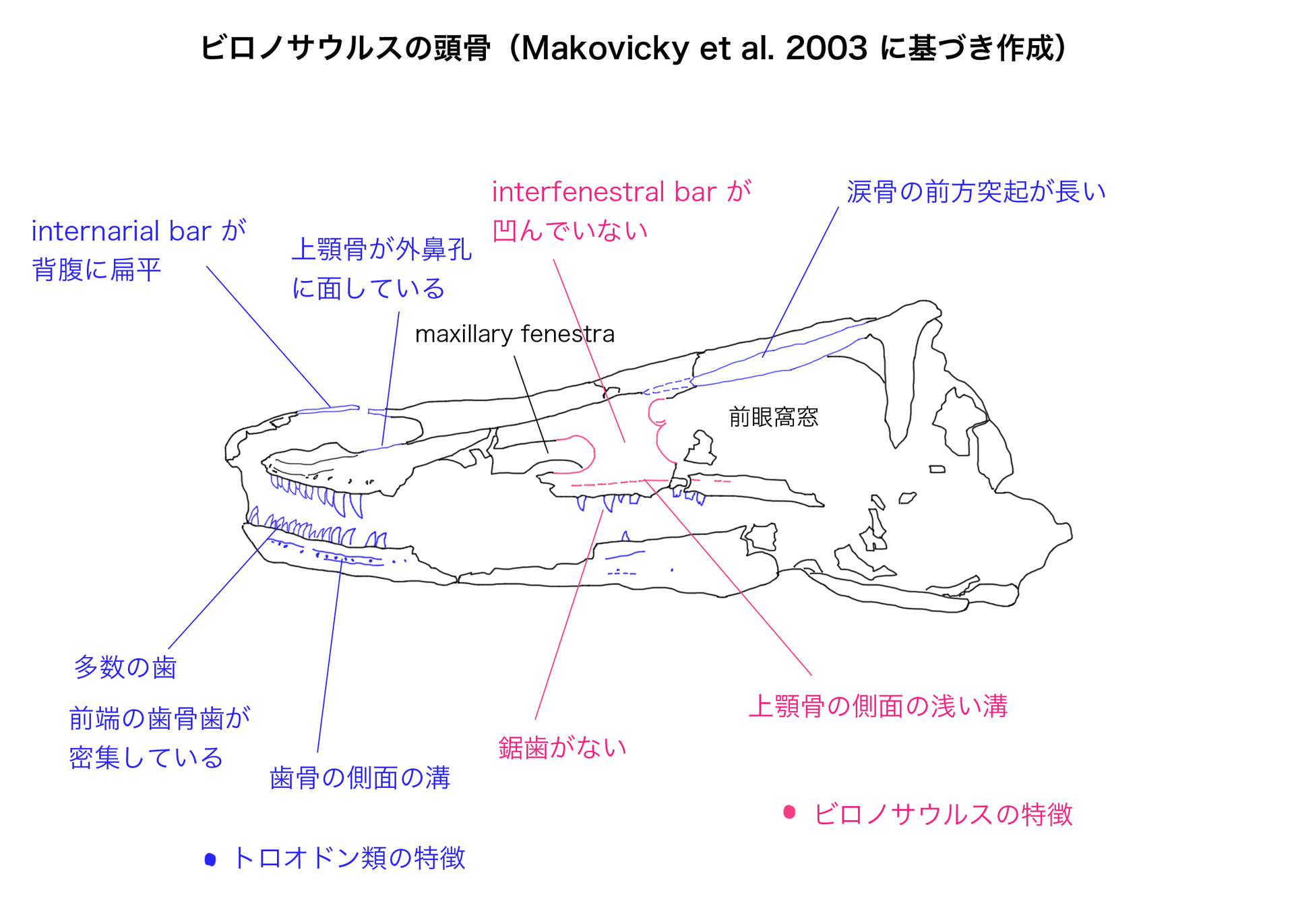
トロオドン類の顔つきを比較しているが、ここで急に時代が飛んで、顔つきも細長くなってしまう。ビロノサウルスはトロオドン類の中でも特に吻が長く、歯の数も最も多いものである。
ビロノサウルスは、白亜紀後期おそらくカンパニアン(Djadokhta Formation)にモンゴルのウハートルゴドに生息した小型のトロオドン類で、2000年に簡単に記載され、2003年のモノグラフで詳細に記載された。鋸歯のないトロオドン類としては最初に発見されたものである。ホロタイプは保存のよい吻部と脳函、いくつかの断片的な体の骨(頸椎、膝関節、趾骨など)からなる。参照標本としていくつかの頭骨の断片(特徴からビロノサウルスと同定)が知られている。
昔はサウロルニトイデスやトロオドンのような派生的なトロオドン類しか知られていなかったため、鉤状の大きな鋸歯のある特徴的な歯の形状がトロオドン類の特徴とされていた。しかしビロノサウルスに続いて、メイ、ウルバコドン、アンキオルニスなど次々に鋸歯のないトロオドン類が発見されたため、大きな鋸歯のある歯は派生的な一部のトロオドン類の共有派生形質と考えられるようになった。
他のトロオドン類と区別されるビロノサウルスの特徴は簡単で、1)歯に鋸歯がないこと、2)interfenestral barが上顎骨の側面から凹んでいないこと(上顎骨の側面と同一平面上にある)、3)上顎骨の側面に歯列と平行な浅い溝があること、である。
Maxillary fenestra と前眼窩窓の間の板状の部分をinterfenestral barという。この用語は、このビロノサウルスの論文(2003)で初めて名付けられたようである。interfenestral barは前眼窩窩の一部なので、通常は上顎骨の表面から凹んだ位置にあるが、ビロノサウルスではこれが上顎骨の表面と同じ平面上にあるのが特徴である。ザナバザル、サウロルニトイデス、シノルニトイデスではinterfenestral barは凹んでいる。
ビロノサウルスにみられるトロオドン類の共有派生形質は、多数の歯、下顎の前端で歯骨歯が密集していること、歯骨の側面に神経血管孔を収める顕著な溝があること、背腹に扁平なinternarial bar、である。
またシノヴェナトル以外のトロオドン類が共有する形質として、脳函に‘‘lateral depression’’という含気性の窪みがある;涙骨の前方突起が長く、前眼窩窓の背側縁をなす;外鼻孔の後縁に上顎骨が面している;を示すという。
歯の数が多いことはトロオドン類の特徴の一つである。ビロノサウルスでは保存されていない部分があるため正確な歯の数はわからないが、前上顎骨歯が4, 上顎骨歯が30, 歯骨歯が30 と推定されている。(他のトロオドン類についてはシシアサウルスの記事参照。)ドロマエオサウルス類では歯の数はもっと少なく、ドロマエオサウルスで上顎骨歯9、歯骨歯11、デイノニクスで上顎骨歯15、歯骨歯16となっている。原始的なオルニトミムス類ペレカニミムス、アルヴァレスサウルス類シュヴウイアやテリジノサウルス類も多数の歯をもつが、これらはトロオドン類とは独立に獲得したと考えられている。
internarial barとは、左右の前上顎骨の鼻骨突起からなる細い部分で、ほとんどの獣脚類では丸い棒状の構造である。トロオドン類ではこれが背腹に扁平になっている。前上顎骨の鼻骨突起は高さよりも幅が広く、背側表面が平坦になっている。これはドロマエオサウルス類やとさかのないオヴィラプトロサウルス類の丸いinternarial barとは対照的である。扁平なinternarial barはオルニトミモサウリアとアルヴァレスサウルス類にもみられ、独立して進化したと考えられている。
細かいことであるが、ビロノサウルスではinterfenestral barの後縁、つまり前眼窩窓の前縁に2つの穴がある。X線CTなどにより内部構造を観察した結果、背側の穴は鼻腔に、腹側の穴はmaxillary fenestraに通じていることがわかった。サウロルニトイデスやザナバザルでも前眼窩窓からmaxillary fenestraに通じる穴があるようだ。するとシヌソナススの論文に「前眼窩窓とmaxillary fenestraをつなぐ通路がない」とあるのは、これらとの違いを意識したものかもしれない。
参考文献
Norell, M.A., Makovicky, P.J., and Clark, J.M. (2000) A new troodontid theropod from Ukhaa Tolgod, Mongolia. Journal of Vertebrate Paleontology 20: 7-11.
Makovicky, P.J., Norell, M.A., Clark, J.M., and Rowe, T. (2003) Osteology and relationships of Byronosaurus jaffei (Theropoda: Troodontidae). American Museum Novitates 3402: 1-32.
コメント ( 0 ) | Trackback ( 0 )
シヌソナススの鼻が気になる
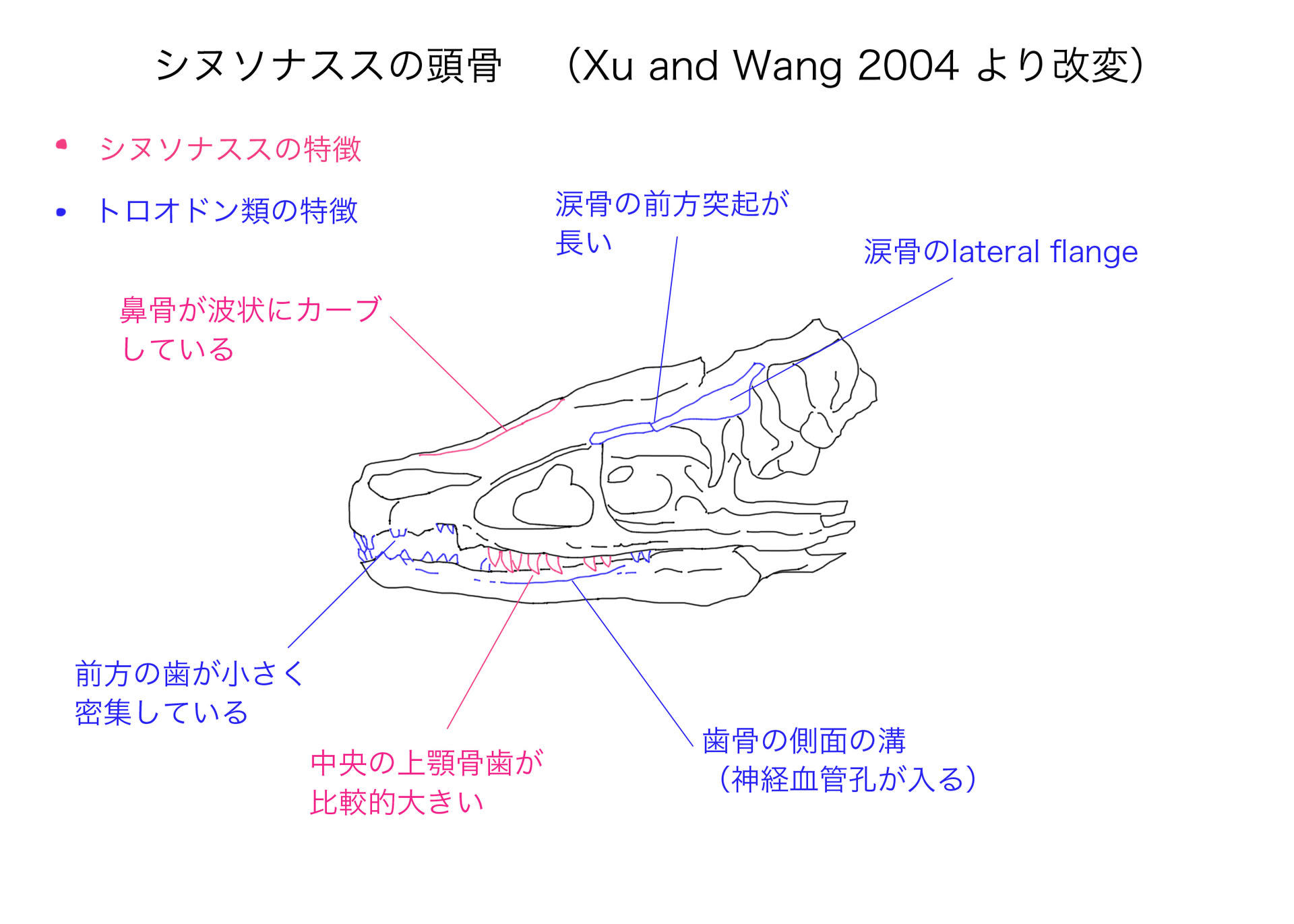
シヌソナススは、白亜紀前期オーテリヴィアン(Yixian Formation)に中国遼寧省に生息した小型のトロオドン類で、2004年に記載された。ホロタイプは仙前椎、肩帯、前肢を除く全身骨格、つまり頭骨と後半身からなる。なかなか見事な化石である。
トロオドン類の顔つきの変遷を確認する上で重要な種類であるが、ちょっと気になる点がある。
シヌソナススの第一の特徴は、鼻骨が側面から見て波状にカーブしていることであるという。しかしまず、記載論文の写真と比べて、線画では鼻骨のカーブがかなり強調されているように見える。また写真では、鼻骨の輪郭はごくゆるやかなS字状にみえる。S字状というなら理解できるが、著者らが本文中で言っていることはそうではない。Xu and Wang (2004) は、「鼻骨は外鼻孔の上で凹んでおり、maxillary fenestraの上で再び凹んでいるので、正弦波状sinusoidの輪郭をなす」と言っているのである。外鼻孔の後半の上で凹んでいるのはわかる。しかしその後方では、鼻骨は額にかけて盛り上がっているだけのように見える。つまりmaxillary fenestraの上では、むしろ盛り上がっているように見える。著者らのいう通りとすればS字状ではなく、W字状ということになるが、そうは見えない。シヌソナススの鼻骨に詳しい方は教えていただきたい。前頭骨あたりの感じから少し斜めにつぶれているように見えるので、鼻骨も斜めになっており、曲面形状としてW字状という意味かもしれない。
それ以外のシヌソナススの特徴は、前眼窩窓とmaxillary fenestraをつなぐ通路がない(詳しい説明はない)、比較的大きな歯、尾の大部分に沿ってプレート状の血道弓が帯状の構造をなす、大腿骨頭と骨幹の間に長い頸状部がある、であるという。
涙骨はT字形で、長い前方突起と、下行突起の上に張り出した顕著な側方の突起(図のlateral flange)がある。他のトロオドン類と同様に、三角形に近い歯骨の側面に、神経血管孔の入る溝がある。ただし溝は比較的浅いという。
図では名称は示していないが、上顎骨には比較的小さい前眼窩窓、大きなmaxillary fenestra、スリット状のpromaxillary fenestraがあるという。maxillary fenestraが大きいので、確かにinterfenestral bar は細くなっている。
上顎骨には約19本の歯がある。上顎骨歯も歯骨歯も、前方では小さく密集しており、後方では大きく間隔が空いているというトロオドン類の特徴を示す。中央の上顎骨歯は最も大きく、他のトロオドン類と比べて比較的大きい。前方の歯には鋸歯がなく、後方の歯の後縁だけに鋸歯がある。それらは比較的小さく、鉤状の先端はない。つまり後の進化したトロオドン類の鋸歯とは異なる。
約30個の尾椎が保存されている。後方の尾椎は、背側の溝sulcusなどのトロオドン類の特徴を示す。シヌソナススの特徴として、中央と後方の血道弓はプレート状で、互いに接して尾椎の腹側に帯状構造を形成している。恥骨は関節していないが、関節面の形状などから前腹方を向いていたと考えられている。坐骨はサウロルニトイデスのような派生的なトロオドン類とよく似ており、大きな三角形の閉鎖突起が中程に位置している。
中足骨は典型的なアルクトメタターサル状態を示す。また他のトロオドン類と同様に、第 II 中足骨は短く細く、第 IV 中足骨は太い。派生的なトロオドン類やドロマエオサウルス類と同様に、第 II 指は高度に特殊化している。
感想としては、植物の実や昆虫をついばむ小鳥のようなメイの顔に比べれば、シヌソナススはやや顎が大きく、小型ながらドロマエオ的な肉食恐竜の顔つきになっている。トカゲくらい捕食してやるという気持ちが感じられる。
早くもアルクトメタターサルとなり、鋸歯があるなど、地上性のハンターとして進化し始めた先駆け的な種類なのだろうか。
参考文献
Xu X, Wang X (2004) A new troodontid (Theropoda: Troodontidae) from the Lower Cretaceous Yixian Formation of Western Liaoning, China. Acta Geol Sin-Engl 78: 22-26.
コメント ( 0 ) | Trackback ( 0 )
眠り竜は二度寝する
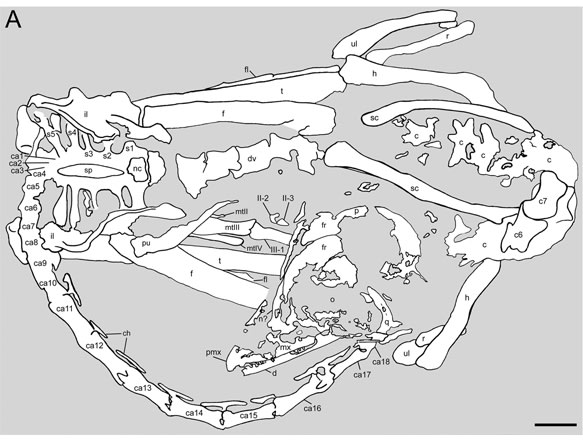
うっかりして1億年の眠りについてしまった寝姿。Copyright 2012 Gao et al.
面白いタイトルを付けたかっただけで、内容はトロオドン類の特徴がメイではどうなっているのか、メイの特徴との関連はどうかということである。
詳しい人はご存知と思うが、中国でメイの第2標本が発見され、この個体もやはりホロタイプと同様に、体を丸めて眠る姿勢で保存されていた。二度寝ではないとしても、非常に稀有なことであり、この動物の生活様式の中で典型的な姿勢であったことは確かである。
この第2標本の発見によってメイの特徴が若干追加された。また後肢の骨の断面を組織学的に観察した結果、体は小さいけれども2才以上の成体と考えられた。新しいデータを追加して系統解析した結果は、従来の解析とあまり変わらず、メイはトロオドン類の中で基盤的な位置にきた。ビロノサウルスやタロスとの関係が多少変わっている。

改訂されたメイの特徴は、外鼻孔が非常に大きく、上顎骨の歯列の1/2 を越えて後方にのびる;中央部の上顎骨歯が密集している;上顎骨の歯列がpreorbital barの位置まで後方にのびている;仙骨の後方部が非常に幅広く、長く延びた第4・第5仙肋骨をもつ;背面からみて腸骨が強くS字状で、側面へのカーブはヴェロキラプトルとアンキオルニスよりも強い;などである。
さらに他の種類にもある形質の組み合わせとして、前頭骨の後方部分が丸く膨らんでいる、短くとがった吻、小さく丈の低いmaxillary fenestraと丈の低い前方突起をもつ上顎骨、後縁がまっすぐな鋸歯のない歯、U字型に近い叉骨、上腕骨の90%以上の長い橈骨と尺骨、中手骨III がIIより長い、などを示す。
Gao et al. (2012) は、この第2標本にみられるトロオドン類の特徴として、歯骨の側面に神経血管孔を収める溝がある、方形骨の後方面に含気孔がある、多数の歯、歯骨と上顎骨の前方の歯が密集している、後方の尾椎には神経棘がなく、代わりに背側正中の溝sulcusがある、をあげている。写真を見る限り、この第2標本では歯列の保存状態はあまりよくないようで、多数の歯とか前方で密集しているというのは難しいようにもみえる。例えばメイの特徴とされる中央の上顎骨歯は線画には描かれていない。確認が難しいということではないだろうか。
一方ホロタイプの方は、上顎骨の歯列がよく保存されている。Xu and Norell (2004)によると、他のトロオドン類と同様に、多数の上顎骨歯(24)があり前方で密集している。さらにメイの特徴として中央の上顎骨歯さえも密集している。後方の歯は太く後方にカーブしている。また歯骨の側面に溝があり、その中に神経血管孔が並んでいる。Internarial barはひも状とある。メイでは多くのトロオドン類と異なりmaxillary fenestraは小さく、interfenestral barという言葉も出てこない。吻が短くとがっているので、maxillary fenestraが大きくなる余地はない感じである。
メイの足はシノヴェナトルと同様に、アルクトメタターサルではない。これらの中足骨は第3中足骨が細くなりアルクトメタターサルに近づいているが(サブアルクトメタターサル)、まだアルクトメタターサルにはなっていない。
メイの第2標本では恥骨は関節してはいないが、保存状態から恥骨は前腹方を向いていたと考えられている。これは後方を向いているシノヴェナトルとは大きく異なる。
9番目より後方の尾椎(10-18)は同じ形をしていて、椎体は長く、側面が平坦またはわずかに凹んでいる。この部分の尾椎には神経棘と横突起がなく、前後に走る背側の溝sulcus がある。この溝は、"サウロルニトイデス"、ビロノサウルス、シノルニトイデス、シヌソナスス、トロオドンにもみられる。
参考文献
Gao C, Morschhauser EM, Varricchio DJ, Liu J, Zhao B (2012) A Second Soundly Sleeping Dragon: New Anatomical Details of the Chinese Troodontid Mei long with Implications for Phylogeny and Taphonomy. PLoS ONE 7(9): e45203. doi:10.1371/journal.pone.0045203
Xu X, Norell MA (2004) A new troodontid dinosaur from China with avian-like sleeping posture. Nature 431: 838-841.
コメント ( 0 ) | Trackback ( 0 )
アンキオルニスはどの辺がトロオドン類?
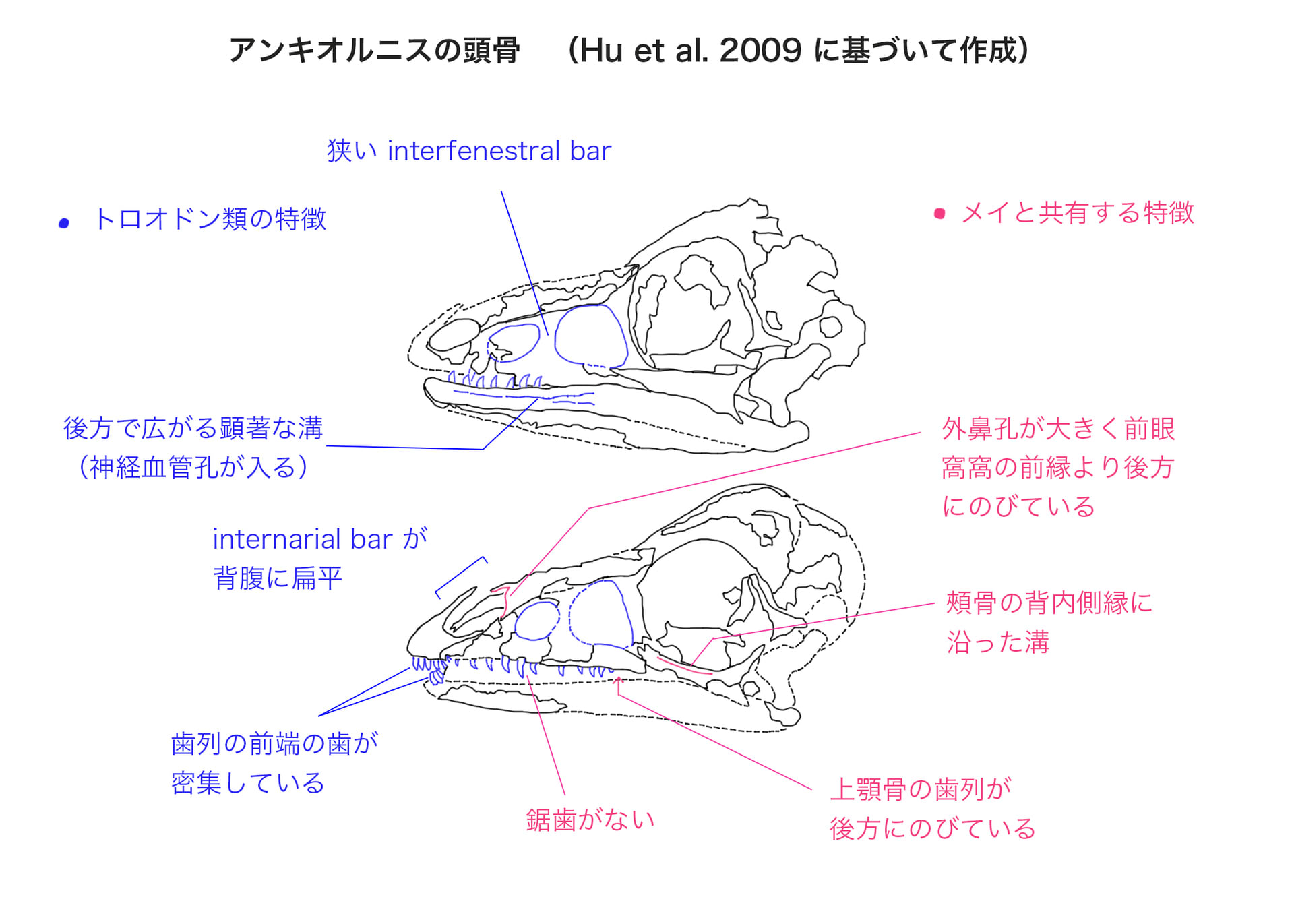
アンキオルニスといえば、今は圧倒的に「羽毛の色」であり、羽毛の構造や進化も話題になるが、骨に注目した紹介は少ない。私は獣脚類の「顔つき」に興味があるので、トロオドン類の特徴がどのように進化してきたのか、簡単に確認したい。
アンキオルニスの段階では、始祖鳥やミクロラプトルなどと大差ない、基盤的なパラヴェス類の顔である。眼窩が大きめの単純な顔つきで、後のトロオドン類のような特徴的な鋸歯もなく、吻が長くもない。これのどの辺がトロオドン類なのだろうか。
アンキオルニスがトロオドン類と共有する形質は、1)大きなmaxillary fenestra と前眼窩窓が狭いinterfenestral barで隔てられている、つまりmaxillary fenestraが拡大しているので、前眼窩窓との間の骨がinterfenestral barとよばれる細い板状の部分だけになっている、2)左右の鼻孔の間のinternarial bar が背腹に扁平である、3)歯骨の側面に後方で広がる顕著な溝があり、その中に神経血管孔が収まっている、4)歯列の前方の前上顎骨歯と歯骨歯が密集している、5)胴椎と前方の尾椎の横突起が細長い、である。
またアンキオルニスは、メイといくつかの形質を共有しており、これもアンキオルニスがトロオドン類に位置づけられる要因となっている。それらは、1)外鼻孔が大きく前眼窩窩の前縁より後方にのびている、2)頬骨の眼窩下突起の背内側縁にそって溝がある、3)鋸歯がない、4)上顎骨の歯列が後方にのびている、などである。
後方の歯列についてHu et al. (2009) は、まばらに分布している点はアヴィアラエに似ているといっている。多数の歯が並んでいるわけではないので、トロオドン類の状態とは異なるということだろう。しかし前方と後方で分布が異なる(前方では密集し、後方では間隔があいている)という意味では、すでに前兆がみられるといってもいいのではないだろうか。つまり歯列の分布と歯骨の溝は、由緒正しいトロオドン類の特徴といえるだろう。また大きなmaxillary fenestraに狭いinterfenestral barというのも、後のビロノサウルスなど吻の長い種類では顕著だが、意外と由緒正しい形質なのだなというのが感想である。
注)Hu et al. (2009) には頭骨復元図も載っているが、ここではそれを用いず実物のスラブ・カウンタースラブの線画を用いた。その理由は、以下である。Hu et al. (2009)はmaxillary fenestraの前方にpromaxillary fenestraがあると考えて描き込んでいるが、写真と線画ではpromaxillary fenestraの形が保存されているようにはみえない。そしてpromaxillary fenestraを描き込んだために、maxillary fenestraが後方にずれてinterfenestral barが実際よりも細くなってしまっている。また実物ではmaxillary fenestraの位置に大きい上顎骨歯が3本あるが、頭骨復元図ではその位置関係がずれている。歯骨の前方と後方の歯の大きさも少し変わっている。
ちなみに、メイと同様に「外鼻孔が大きく前眼窩窩の前縁より後方にのびている」とあるが、「前眼窩窩の前縁」は破損した部分にあるようにみえる。明確に見えているのではなく推定ということだろう。
参考文献
Hu, D. Y., Hou, L. H., Zhang, L. J. & Xu, X. (2009) A pre-Archaeopteryx troodontid from China with long feathers on the metatarsus. Nature 461, 640-643.
コメント ( 0 ) | Trackback ( 0 )
シシアサウルス
大きい画像
シシアサウルスは、白亜紀後期コニアシアンからカンパニアン(Majiacun Formation)に中国河南省の西峡盆地Xixia Basinに生息したトロオドン類で、2010年に記載された。モンゴルのビロノサウルスやウズベキスタンのウルバコドンに続く、歯に鋸歯のないトロオドン類で、ビロノサウルスと最も近縁と考えられている。
ホロタイプ標本は部分的な頭蓋、下顎の前方部分、橈骨・尺骨の断片(中央部)、完全な第I指を含む部分的な右手からなる。(ここでは手の指はI, II, II)
他のトロオドン類と区別されるシシアサウルスの特徴としては、上顎骨歯が22本であることがあげられる。これはビロノサウルス(30以上)やメイ(24)よりも少なく、シノルニトイデス(18)、ザナバザル(20)、サウロルニトイデス(19)よりは多い。また、前上顎骨の鼻骨突起の基部に顕著な孔があること、上顎の前端がビロノサウルスよりも先細りのU字形をしていること、歯骨の先端の顎間結合の部分がわずかに内側に曲がっていることもあげられる。
ビロノサウルス、メイ、ウルバコドン、アンキオルニスと同様に、シシアサウルスの歯には鋸歯がない。またビロノサウルスと同様に長く延びた二次口蓋をもつ。
Lu et al. (2010) によると、シシアサウルスは以下のような形質に基づいて、トロオドン類に分類される。他の多くの獣脚類よりも歯の数が多いこと、歯冠と歯根の間がくびれていること、下顎の前端で歯骨歯が密集していること、歯骨の側面に神経血管孔を容れる顕著な溝があること、である。歯列については上顎でも同じ傾向があるので、不均一な分布と異歯性といってもいいだろう。歯骨の側面の溝については、論文の写真では他のトロオドン類ほどくっきりしてはいないように見えるが、保存の問題かもしれない。また歯骨の前端部では個々の歯槽が癒合して、ひとつながりの溝になっていることも記載している。
アジアでは多数のトロオドン類が報告されているが、歯の形態は非常に多様である。もともと白亜紀後期のトロオドン類では、多数の小さい歯と特徴的な大きい鋸歯が知られていた。白亜紀前期のシノヴェナトルの歯はドロマエオサウルス類のような小さい鋸歯をもつので、これが祖先形と考えられた。ところがジュラ紀後期のアンキオルニスの発見により、これが最古のトロオドン類となってきた。アンキオルニスの歯は鋸歯がないので、トロオドン類の祖先が鋸歯を失うという変化はさらに古く、ジュラ紀前期に起きたことになる。そして白亜紀後期のトロオドン類には大きな鋸歯をもつものと、鋸歯をもたないものがいるというように、トロオドン類の歯は非常に複雑な歴史をもつことがわかってきたようである。
ビロノサウルス、ウルバコドン、シシアサウルスの歯に鋸歯がないことは、おそらく食性の変化と関係しており、植物食あるいは雑食だったかもしれないといっている。ただし、あまり詳しい議論はしていない。トロオドン類の大きな鋸歯自体がもともと、肉食にも植物食にも適しているという議論があったのではなかったか。鋸歯を失うイコール植物食という論法にはちょっと引っかかるものを感じる。鋸歯を失っただけでは、大型動物の肉を切り裂くには適さないというだけで、積極的に植物食に適応していることにはならないように思える。
タカがハトを解体して食べる際は、くちばしで肉を引き裂いて飲み込んでいる。それに比べれば、シシアサウルスの歯は鋸歯こそないが、鋭い稜縁があり、後方の歯は強く後方に反っている。顎に並んだ多数の歯自体が、全体としてのこぎりの機能を果たしうるのではないか。丸のみできる、あるいは少し引き裂けば飲み込めるくらいの小動物を捕食するのには、鋸歯がなくても大して困らなかったということはないのだろうか。スピノサウルス類は鋸歯を失ったが、魚などの小動物を捕食するためとされている。鋸歯のあるトロオドン類に比べて、より小型の獲物を捕食したということならわかる。
また歯列が異歯性であることからトロオドン類全体として雑食性で、白亜紀後期の大きな鋸歯をもつ種類は肉食傾向が強い(肉食に特化していた)、ということかもしれない。鋸歯のないものはより雑食性で、動物質は昆虫やトカゲくらい、ということか。
参考文献
Lu, J.-C., Xu, L., Liu, Y.-Q., Zhang, X.-L., Jia, S.H., and Ji, Q. (2010) A new troodontid theropod from the Late Cretaceous of central China, and the radiation of Asian troodontids. Acta Palaeontologica Polonica 55 (3): 381-388.
(Luはウムラウト)
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ |