獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年9月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ブイトレラプトル


大きい画像
ブイトレラプトルは、白亜紀後期セノマニアンからチューロニアン(Candeleros Formation)にアルゼンチンのリオ・ネグロ州Rio Negro Provinceに生息したドロマエオサウルス類で、2005年に記載された。ゴンドワナに分布したウネンラギア類の中では、最も標本が完全で、最も時代が古いものとされる。というより、Makovicky et al. (2005)によるブイトレラプトルの研究によって、ゴンドワナのドロマエオサウルス類が単系群と認識され、それがウネンラギア亜科 Unenlagiinae と命名された。
ホロタイプには全身のかなりの部分の骨が含まれているが、多くの骨が部分的に失われている。フィールド博物館など海外には復元骨格があるようだが、日本に来たことはないと思われる。
Turner et al. (2012) によると、ブイトレラプトルは次の形質の組み合わせと固有形質により識別される小型のドロマエオサウルス類である。これらのほとんどはMakovicky et al. (2005) と同じである。頭骨が長く、大腿骨の長さよりも25%大きい;歯は小さく、鋸歯がなく、歯冠と歯根の間がくびれていない;方形骨には大きな側方フランジlateral flangeと含気孔がある;後方の頸椎の椎体に腹方外側の稜がある、などである。また他のドロマエオサウルス類と区別される形質として、maxillary fenestra が大きい;前頭骨の縁から後眼窩骨突起までが連続的に移行している(トロオドン類と共有);歯骨に深い歯槽下溝subalveolar groove がある(トロオドン類と共有)、などを追加している。
頭骨はある程度の骨が見つかっているものの、欠損部分が多いので全体の形は推定で復元されている。顔が非常に細長いのは確かで、トロオドン類を通り越して翼竜のような印象もある。一方、歯のついた上顎骨と歯骨は保存されているので、ブイトレラプトルの歯について研究した論文が出ている。
ブイトレラプトルの歯は頭骨のわりに小さく、歯冠の高さが0.6 から 4.6 mmである。歯は後方にカーブし、扁平で、先が非常に尖っている。歯槽に保存された歯も分離した歯も、大きさが異なるだけでみな同じ形をしている。左の歯骨に保存された歯の一つは歯槽の縁が欠けていて、それをみると歯冠と歯根の間が連続的でくびれていない。この点はトロオドン類などと異なる。すべての歯において歯冠の前縁も後縁も少し丸みを帯びていて、鋸歯も稜縁もない。
歯冠の外側(唇側)と内側(舌側)は非常によく似ていて、曲がりがないので唇側と舌側を区別することは困難である。重要な特徴として唇側と舌側の両面に縦のくぼみdepressionがあるため、歯冠の断面が8の字形になっている。さらに、両面に平行に並んだ縦の溝と稜がある。これらの溝と稜は一定のパターンをもつものではなく、数も一定しない。たとえば2本の溝がある場合は稜、溝、稜、溝、稜と並んでいる。
ドロマエオサウルス類の歯の特徴は、より多くの種類が知られるローラシアのドロマエオサウルス類について研究されてきたが、ゴンドワナのウネンラギア類(ブイトレラプトルとアウストロラプトル)は、ローラシアのドロマエオサウルス類とはかなり異なった歯の形態を示すことがわかってきた。Gianechini et al. (2011) は、ウネンラギア類の歯の特徴として1)歯の数が多い、2)歯のサイズが小さい、3)鋸歯も稜縁もない、4)歯冠の表面に縦の溝grooveがある、を挙げている。
1)歯の数が多い:ローラシアのドロマエオサウルス類では下顎の歯骨に11 から16本の歯があるが、ブイトレラプトルでは約25本と推定されている。アウストロラプトルも、少なくとも25本と考えられている。また多くのドロマエオサウルス類は9 から15本の上顎骨歯をもつが、アウストロラプトルは24本である。このような多数の歯はトロオドン類やオルニトミモサウリアのペレカニミムスなどにもみられる。
2)歯のサイズが小さい:歯の相対的な大きさは、上顎骨の最も大きい歯と、上顎骨の丈の高さ(前眼窩窓の前縁の位置で)の比率で表される。この比率はローラシアのドロマエオサウルス類では0.25-0.40 であるが、ブイトレラプトルでは0.18 である。系統解析の結果からするとウネンラギア類の小さい歯は、ローラシアのドロマエオサウルス類にみられる大きい歯が縮小したものと考えられるという。より広くマニラプトル形類をみると、トロオドン類やアヴィアラエ類が0.20より小さく、またテリジノサウルス類エルリコサウルスやアルヴァレスサウルス類シュヴウイアも小さいことから、広く分布する収斂と考えられる。
3)鋸歯も稜縁もない:ローラシアのドロマエオサウルス類では一般に歯の前縁と後縁に鋸歯があるが、ウネンラギア類では前縁にも後縁にも全く鋸歯がない。この鋸歯の分布についてはちょっと複雑である。
ドロマエオサウルスでは前縁にも後縁にも同じくらいの大きさの鋸歯がある。ヴェロキラプトルとデイノニクスでは、前縁の鋸歯が後縁の鋸歯よりずっと小さい。サウロルニトレステスも同様だが、この種では前縁の鋸歯があるものとないものがみられる。ミクロラプトルの前上顎骨歯と前方の歯骨歯には鋸歯がない。シノルニトサウルスでは、前上顎骨歯、最初の上顎骨歯、前方の歯骨歯には全く鋸歯がなく、その後方の一部の歯では後縁だけに鋸歯がある。ドロマエオサウルス類では、一部の歯で前縁の鋸歯を失っているのが基本で、その後二次的に前縁の鋸歯を回復してきたと考えられているらしい。(それでドロマエオサウルス亜科が派生的となるようである。)その中にあって、完全に鋸歯がないことはウネンラギア類の共有派生形質と考えられるという。
4)歯冠の表面に縦の溝grooveがある:ウネンラギア類の特徴の一つは歯冠に縦の溝があることである。この溝は、系統的には離れているがスピノサウルス類やケラトサウルスにみられるものと若干似ている。El Anfiteatroという場所で発見された分離した歯、Endemas-Pv 15はウネンラギア類と比較されるマニラプトル形類とされている。この歯は鋸歯がなく、強く後方に曲がっており、断面が8の字形であり、またエナメルのひだfoldがある点で、ブイトレラプトルと似ている。ただし大きさがブイトレラプトルよりずっと大きく、19 mm もあることから、別のウネンラギア類と考えられる。他にもドロマエオサウルス類の可能性がある歯で、溝や稜のあるものがモロッコ、スペイン、フランスなどから報告されているが、これらは鋸歯があるという。
Gianechini et al. (2011) は、縦の溝のある歯はスピノサウルス類、ノアサウルス類、モササウルス類、翼竜など、魚食性の四足動物にみられることは興味深いといっている。ブイトレラプトルもアウストロラプトルも、豊富な魚類化石を含む河川堆積物の近くで発見されていることから、魚食性の可能性があると述べている。このスコミムスか偽歯鳥類のような長い吻を、水面下に突っ込んで魚類などを捕食したのだろうか。
これほど特殊化した動物が、ドロマエオサウルス類の中で基盤的な位置にくる理由は何だろう。Makovicky et al. (2005) によると肩帯、腰帯、後肢などに系統上重要な形質があるようだ。烏口骨は直角に曲がっているが、これはシノルニトサウルスと似ている。上腕骨は肩甲骨よりも30%長いが、これは鳥類と遼寧省のドロマエオサウルス類(ミクロラプトル類)にしかみられない。腸骨には顕著なsupra-acetabular crestがあり、これはウネンラギアやラホナヴィスと同様である。座骨の閉鎖突起obturator processの形は、ラホナヴィス、ミクロラプトル、シノルニトサウルスと似ている。中足骨はアルクトメタターサルで、尾椎の前関節突起はあまり長くない。これらはドロマエオサウルス類としては原始的な形質ということかもしれない。
参考文献
Gianechini, F.A., Makovicky, P.J., and Apesteguia, S. (2011). The teeth of the unenlagiine theropod Buitreraptor from the Cretaceous of Patagonia, Argentina, and the unusual dentition of the Gondwanan dromaeosaurids. Acta Palaeontologica Polonica 56 (2): 279-290.
Makovicky, P.J., Apesteguia, S., and Agnolin, F.L. (2005). The earliest dromaeosaurid theropod from South America. Nature 437: 1007-1011.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )
ドロマエオサウルス補足
ドロマエオサウルスの前上顎骨歯について、前の記事中の文言を一部訂正し、ここに補足説明します。
ヴェロキラプトルでは2番目の歯が最も大きいのに対して、ドロマエオサウルスでは4本とも同じくらいとされる理由は、分離した前上顎骨歯が見つかっているため。
Colbert and Russell (1969) によると、歯槽に保存されているのは左の前上顎骨の後半部分の2本(3番と4番)だけであるが、その他に4本の分離した前上顎骨歯が見つかっている。形態から、これらのうち2本は右側、2本は左側のものである。さらに前縁と後縁の近接の程度(D字形に近い)から、右の1本と左の1本は1番目の前上顎骨歯と考えられる。よって左側の残りの1本は2番目となり、左側は一応、1、2、3、4が揃っていることになる。そのためCurrie (1995)の論文中で、大きさが4本とも同じくらいとされている。それなりのロジックがあって結論されているのが面白い。
ドロマエオサウルスの最大の特徴である、歯の前縁のねじれを確認しようと思って、Colbert and Russell (1969)を参照したのであるが、なんと前上顎骨歯の写真はあるが、上顎骨歯と歯骨歯の写真はないようだ。上顎骨歯については、文章で記述されている。
参考文献
Colbert, E.H. and Russel, D.A. (1969). The small cretaceous dinosaur Dromaeosaurus. American Museum Novitates 2380: 1-49.
コメント ( 0 ) | Trackback ( 0 )
ドロマエオサウルス


大きい画像
ドロマエオサウルスは、白亜紀後期カンパニア期(Oldman Formation)にカナダのアルバータ州に生息した中型のドロマエオサウルス類で、Matthew and Brown (1922) によって記載された。Matthew and Brown (1922)の記載は、部分的にクリーニングされた標本に基づく予備的なものだったが、その後Colbert and Russell (1969) によってより完全な記載がなされた。さらにCurrie (1995)は、ホロタイプ標本の再クリーニングとCTスキャンによって、頭蓋と脳函の形態について新しい情報を追加し、頭骨の復元を行った。
ホロタイプAMNH FARB 5356 には、部分的な頭骨と下顎、舌骨、足の骨、左の第1中手骨が含まれている。
ホロタイプの頭骨は、上顎、下顎、後頭部の骨が主で、頭蓋天井の大部分は見つかっていない。つまり鼻骨、上顎骨の上部、涙骨のほとんど、後眼窩骨などは保存されていない。前頭骨と脳函はある。

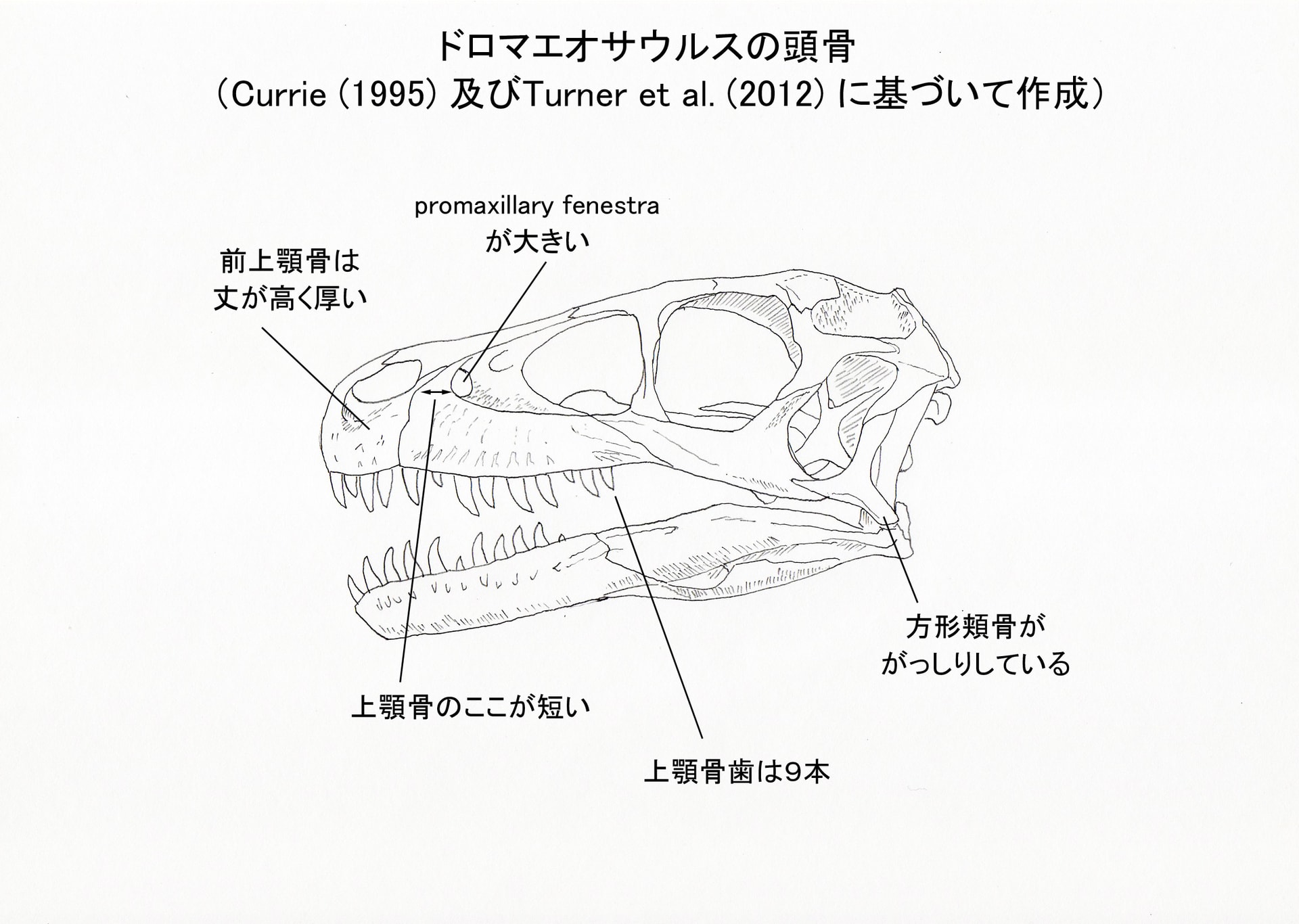
大きい画像
Turner et al. (2012) によると、ドロマエオサウルスは以下の形質の組み合わせと固有形質により識別される中型のドロマエオサウルス類であるという(これらの大部分はCurrie (1995) からそのまま採用している)。ここでは主なものを骨ごとにまとめてみる。
歯:9本の上顎骨歯;上顎骨歯と歯骨歯の前縁anterior carinaが(先端付近では正中にあるが)舌側へねじれている;前縁の鋸歯と後縁の鋸歯の大きさがほぼ同じくらいである。
前上顎骨:(他のドロマエオサウルス類よりも)前上顎骨の丈が高く厚い。
上顎骨:上顎骨の前眼窩窩より前方の部分が前後に短い;拡大したpromaxillary fenestra (または極端に前腹方にあるmaxillary fenestra)。
方形頬骨:方形頬骨はがっしりしているstout。
前頭骨:前頭骨の先端が扁平である;前頭骨の後眼窩骨突起と眼窩の背側縁の境界がはっきりしている;鼻骨と前頭骨の間の縫合線がV字形;前頭骨の前方に涙骨と関節するための深い切れ込みnotchがある。
その他、口蓋骨の形質が1つ、脳函の形質が3つある。ドロマエオサウルスは他のドロマエオサウルス類と異なり、脳函がほとんど含気化していないという。


大きい画像
前上顎骨は、2本の歯のついた後半部分だけが保存されているが、他のドロマエオサウルス類と同様に4本の歯があったと考えられる。ヴェロキラプトルなどでは前上顎骨の2番目の歯が長さ・直径とも最も大きいが、ドロマエオサウルスでは4本とも同じくらいの大きさと考えられる(分離した前上顎骨歯が見つかっている)。
左の上顎骨に、9個の歯槽がある。デイノニクスでは15個、ヴェロキラプトルでは10個、サウロルニトレステスでは少なくとも10個あるという。
涙骨はほとんど失われているが、右の頬骨に付着した小さな断片と、いくつかの分離した断片がある。その形状から、ドロマエオサウルスにはデイノニクスなどにみられるような、眼窩の前背方で側方に突出した膨らみがあったと思われるという。
方形頬骨はT字形で、ヴェロキラプトルなどと似ているが、デイノニクス、ヴェロキラプトル、サウロルニトレステスのものよりもがっしりしている。鱗状骨は、顕著に側方にのびた腹側方突起ventrolateral process をもつなど、ドロマエオサウルス類の特徴を示している。
Turner et al. (2012) の論文中にもドロマエオサウルスの頭骨復元図があるが、ちょっと長めのツァーガン的な顔になっている。これをよくみると疑問な点がある。「上顎骨の前眼窩窩より前方の部分が前後に短い」はCurrie (1995) にはないので、Turnerらが追加した特徴と思われるが、この通りになっていない。図では前眼窩窩より前方の部分が妙に長くなっている。この点で、本文の記述と頭骨復元図が一致していない。Currie (1995)の頭骨復元図の方が信頼できるように思われた。
Turner et al. (2012) の系統解析では、ドロマエオサウルス、ユタラプトル、アキロバトル、アトロキラプトルがドロマエオサウルス亜科Dromaeosaurinae に含まれた。
ヴェロキラプトルなどと比べると、ドロマエオサウルス亜科はどれも不完全である。ユタラプトルやアキロバトルの姿を描こうとすればドロマエオサウルスが参考になるはずだが、ドロマエオサウルス自体が不完全という感じである。ヴェロキラプトルなどよりも上顎、下顎ががっしりしているので、より大型の獲物に挑みそうな、獰猛な印象はある。現代でも生存できそうな洗練されたハンターと考えたい。
参考文献
Currie, P.J. (1995). New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal of Vertebrate Paleontology 15: 576-591.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )
アダサウルスの特徴と系統的位置

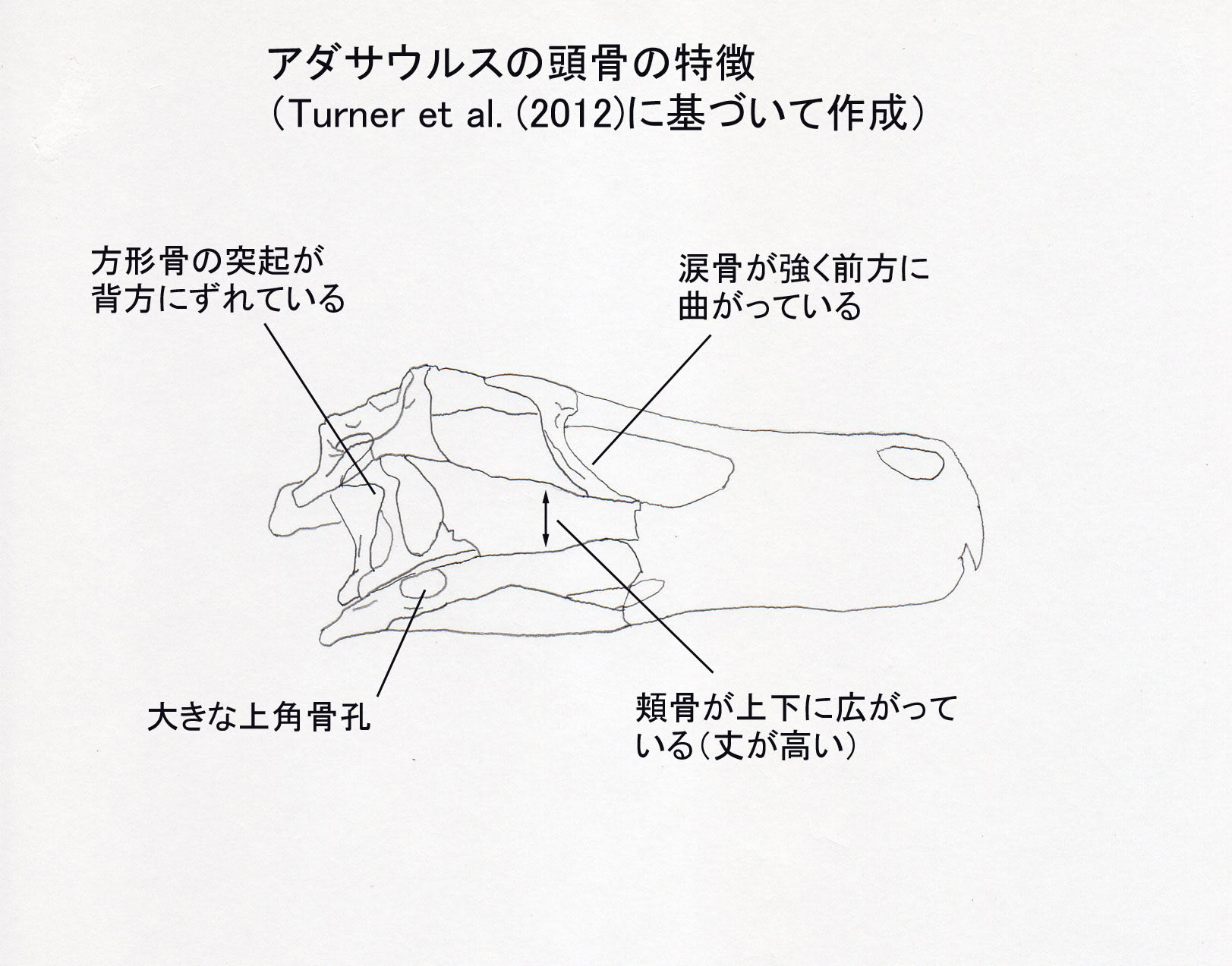
大きい画像
アダサウルスは、白亜紀後期カンパニア期ないしマーストリヒト期(ネメグト層)にモンゴルに生息したドロマエオサウルス類で、Barsbold (1983) によって記載された。ホロタイプIGM 100/20 には頭骨の後半部、脊椎、肩帯、腰帯、後肢の骨が含まれる。パラタイプIGM 100/21 は2個の尾椎と完全な足を含む右後肢からなる。Barsbold (1983)の記載は不十分で、特徴としては後肢の第2指のカギ爪が小さいことしか記述していなかった。またドロマエオサウルス類の中でのアダサウルスの系統的位置ははっきりしていなかった。
アダサウルスの標本は、日本人研究者の久保田克博博士の研究によって再検討された(Kubota and Barsbold, 2006; 2007)。これによりバヤンシレ層から発掘された第3、第4標本(IGM 100/22, IGM 100/23)はアダサウルスではなく、新種のドロマエオサウルス類と考えられた。
Kubota and Barsbold (2006) は、肩甲烏口骨が癒合している点と遠位の足根骨が中足骨と癒合している点で、アダサウルスがヴェロキラプトルと似ていることを見いだした。また他のドロマエオサウルス類と識別できるアダサウルスの特徴として、前頭骨の正中線上に低い稜があること、大きな上角骨孔、腸骨の前寛骨臼突起の前縁に切れ込みがあるnotchedこと、足の第2指のカギ爪が縮小していることを挙げている。ただしこれらの研究はSVPのアブストラクト(学会発表要旨)があるのみで論文にはなっていない。
その後前述のTurner et al. (2012) は、大規模なドロマエオサウルス類の系統研究の中で、アダサウルスのホロタイプ標本を自分で詳細に観察し、多数の新しい形質データをとっている。
Turner et al. (2012) によると、アダサウルスは以下の形質の組み合わせと固有形質により識別される中型のドロマエオサウルス類である。頬骨の上顎骨突起が上下に拡がっている;涙骨の下方突起(腹側突起)が強く前方に曲がっている;大きな上角骨孔;方形骨の側面にある三角形の突起が背方にずれている;前方の仙椎だけにプレウロシール(側腔)がある;腸骨の前寛骨臼突起の前縁に切れ込みがある;足の第2指のカギ爪が縮小している。
ホロタイプの頭骨は、前半部分、つまり吻部が欠けている。また右側はよく保存されているが、左側は保存がよくない。方形頬骨あたりが欠けているので、方形骨がよく見えている。他のドロマエオサウルス類と同様に、方形骨の側面に大きな三角形の突起がある。この三角形の突起が、他のドロマエオサウルス類では中央あたりにあるが、アダサウルスでは背方にずれているという。涙骨の下方突起が強く前方にカーブしているのは、アダサウルスとアウストロラプトルだけに見られる特徴であるという。Kubota and Barsbold (2006)の挙げた前頭骨の低い稜については、Turner et al. (2012) は、前頭骨の後方部分はプレパレーションが不完全で薄い母岩の層で覆われているため確認できないといっている。
方形骨や下顎の孔は生体復元では見えないので、ヴェロキラプトルやツァーガンと描き分けるのであれば、涙骨のカーブが表現しやすいのではないだろうか。(それも普通には見えないが、色を付けたりして)

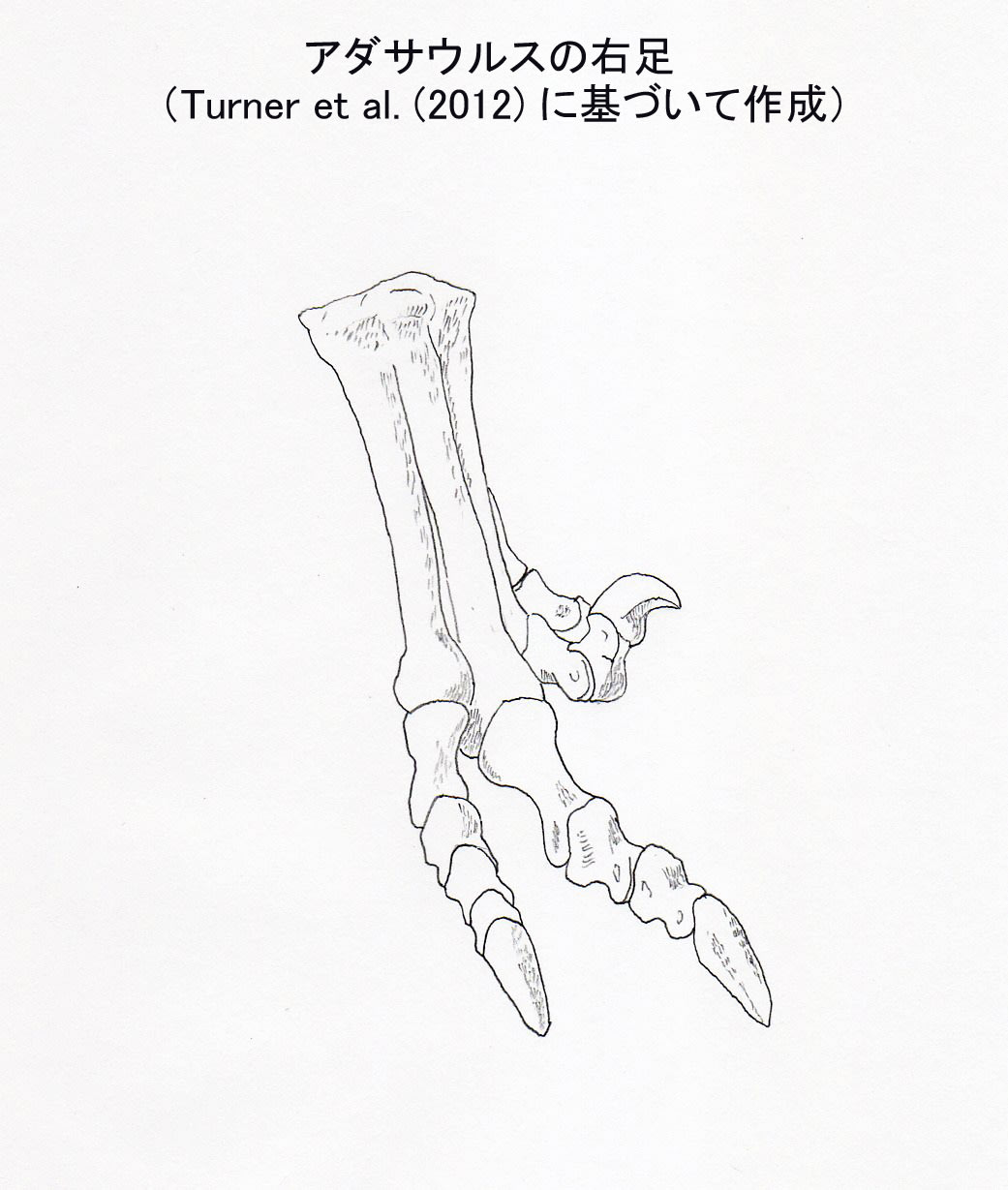
大きい画像
アダサウルスの足の第2指のカギ爪は、他の指のカギ爪と同じくらい小さくなっている。「それでもドロマエオかよ」とトロオドンに突っ込まれそうなレベルである。もはや持ち上げなくてもいいような。。。
少し古い恐竜本には、ドロマエオサウルス亜科とヴェロキラプトル亜科の鋸歯の違いが説明してある。Currie (1995) によると、ヴェロキラプトル亜科の特徴は、上顎骨歯と歯骨歯の前縁の鋸歯が後縁の鋸歯よりも顕著に小さいこと、前上顎骨の2番目の歯が3番目と4番目の歯よりも顕著に大きいこと、であった。これはヴェロキラプトルの頭骨を思い浮かべれば比較的わかりやすいと思える。
しかし2001から2003年ごろの系統解析では、ドロマエオサウルス科の内部の系統関係についてほとんどコンセンサスが得られなかった。よってヴェロキラプトル亜科が本当に単系群であるのか、よくわからなかった。その後、Novas and Pol (2005) など、2005から2007年ごろの系統解析によって、ヴェロキラプトル亜科がクレードとして見いだされた。これにはヴェロキラプトル、デイノニクス、ツァーガンが含まれていた。しかしサウロルニトレステスの位置ははっきりせず、ヴェロキラプトル亜科に含まれたり含まれなかったりした。また、このころはアダサウルスはドロマエオサウルス亜科に含まれていた。
Turner et al. (2012) の大規模な系統解析では、ドロマエオサウルス科の中にウネンラギア亜科、ミクロラプトル亜科、ドロマエオサウルス亜科、ヴェロキラプトル亜科に相当するクレードが見いだされた。彼らはアダサウルスの標本を再検討した結果、102箇所もの形質状態を変更した。これによって、アダサウルスは従来のドロマエオサウルス亜科ではなく、ヴェロキラプトル亜科に分類し直された。
Turner et al. (2012) によると、ヴェロキラプトル亜科にはバンビラプトル、ツァーガン、サウロルニトレステス、デイノニクス、ヴェロキラプトル、アダサウルス、バラウルが含まれた。最後の4種は一つのクレードをなしている。
ではアダサウルスのどこがヴェロキラプトル亜科なのかというと、素人にわかりやすいものではないようだ。ヴェロキラプトル亜科は3つの共有派生形質で支持される。基底蝶形骨の後方の開口部が2つの小さな孔に分かれている、dorsal tympanic recessが深く後側方を向いた凹みである、すべての胴椎にプレウロシールがある、ということである。
デイノニクス、ヴェロキラプトル、アダサウルス、バラウルのクレードの共有派生形質は、第 IV 中足骨の骨幹の断面が幅広く扁平であることであるという。
バラウルを除くと、ヴェロキラプトルとアダサウルスは姉妹群をなした。肩甲烏口骨が癒合していること、遠位の足根骨が中足骨と癒合していること、距骨と踵骨が互いに癒合しているが脛骨とは癒合していないこと、の3つの形質で支持されるという。ただし、これらはいずれも骨同士の癒合に関する形質で、成長段階と関連している可能性もある。そこを明らかにするためには、アダサウルスの追加の標本も含めて更なる研究が必要であるという。
参考文献
Kubota, K., and R. Barsbold. (2006) Reexamination of Adasaurus mongoliensis (Dinosauria: Theropoda) from the Upper Cretaceous Nemegt Formation of Mongolia. Journal of Vertebrate Paleontology 26 (suppl. to 3): 88A.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )
ゼニュアンロン(ジェンユエンロン)


大きい画像
ゼニュアンロン(ジェンユエンロン、ジェニュアンロン、ジェンユアンロン)検索のための文字列
(またやっかいな中国語名で、多分ジェンユエンロンが近いと思うが、きりがないので今回ローマ字読みで妥協する。ティアンユーもティエンユーとかティアニュとか好きに読んで下さい。)
ゼニュアンロンは、白亜紀前期に中国遼寧省に生息したドロマエオサウルス類で、2015年に記載された。羽毛の痕跡を含む、ほぼ全身の骨格が発見されている。ホロタイプはかなり成熟に近い亜成体とされている。
中国遼寧省のドロマエオサウルス類としては、これまでに5つの属が知られていた。チャンギュラプトル、グラキリラプトル、ミクロラプトル、シノルニトサウルス、ティアンユーラプトルである。これらのほとんどは小型の動物で、長い前肢と羽板のある羽毛でできた大きな翼をもっている。唯一ティアンユーラプトルだけは、全長2m近い大型種で、後肢に比べて短い前肢をもっていた。しかし残念ながらティアンユーラプトルの標本には羽毛が保存されていなかったので、このような短い前肢をもつ種類が小型種と同じような大きな翼を持っていたかどうかは、わかっていなかった。今回Lu: and Brusatte (2015) は大型で前肢の短い2番目のドロマエオサウルス類を発見し、新属新種ゼニュアンロン・スニとして記載している。ゼニュアンロンでは羽毛がよく保存されており、小型で前肢の長い種類と同様に、大きな翼と長い尾羽をもつことがわかった。ただし、後肢にはおそらく長い羽毛はないという。
ゼニュアンロンは遼寧省のドロマエオサウルス類としては大型のもので、保存された標本の全長は126.6 cmあり、尾の半分が失われていると考えられるので、実際の全長は165 cmに達すると推定される。これは遼寧省のドロマエオサウルス類の中ではティアンユーラプトルに次いで2番目に大きい。
ゼニュアンロンは、以下の形質の固有の組み合わせをもつドロマエオサウルス類である。撓骨が非常に細く、その骨幹は指骨 I -1 よりも細い;第II中手骨の長さが、第 I 中手骨と指骨 I -1 を合わせた長さよりも短い(他の遼寧省のドロマエオサウルス類では第II中手骨の方が長い);6個の仙椎;前肢が短く後肢のおよそ1/2の長さで、上腕骨/大腿骨の比率が0.65以下、尺骨/大腿骨の比率が0.55以下、手/大腿骨の比率が0.90以下である(この形質は遼寧省のドロマエオサウルス類の中ではティアンユーラプトルとのみ共有する)、などである。
ゼニュアンロンは、前眼窩窩の腹側が鋭い縁で縁取られている(シノルニトサウルスと同様)、腸骨の後寛骨臼突起の後端が尖っている、などの形質でティアンユーラプトルとは異なっている。またゼニュアンロンとティアンユーラプトルでは四肢の比率も多少異なっており、ゼニュアンロンの方が後肢と比べて前肢がより短く(0.48 と0.53)、大腿骨に対して手がより短い(0.76と0.86)。しかし1個体ずつで比較しているだけなので、同種内の変異についてもっとデータがないと差があるとはいえない。このくらいの比率の違いは、ミクロラプトルなど多くの標本がある種類の種内変異の範囲内なので、ゼニュアンロンとティアンユーラプトルを区別する特徴とはいえないとしている。
羽毛は体のいくつかの部位、特に前肢と尾でよく保存されている。前肢の大きな翼は、羽軸と羽枝のある大羽で形成されている。保存上のゆがみにより翼全体の形ははっきりしないが、かなり面積が大きいものである。右の翼では、雨覆、初列風切、次列風切が確認できる。翼の大きさ、形、羽毛の構造は全般的にミクロラプトル、チャンギュラプトル、アンキオルニス、エオシノプテリクスなどに似ている。
右の翼には、約30本の小さい羽毛が尺骨と第III 中手骨に付着しているのが保存されている。これらは、初列風切と次列風切の背側を覆う大雨覆である。多くは尺骨にほぼ垂直に付いているが、第III 中手骨に付いた遠位のものは斜めを向いており、手の長軸に対してほぼ平行になっている。これらの雨覆は現生鳥類のように短く、アルカエオプテリクスで推定されているように長く伸びてはいない。
初列風切と次列風切も確認できるが、雨覆ほど保存が良くはない。これらの羽毛を数えるのは難しいが、約10本の初列風切と20本の次列風切があるようである。長さを測るのも難しいが、初列風切も次列風切も上腕骨の2倍以上の長さがある。これはミクロラプトルや現生鳥類と同じである。アンキオルニスやエオシノプテリクスでは1.5倍であるという。保存の良い右の翼では初列風切の方が次列風切よりも長い。次列風切は尺骨に対して垂直方向を向いているが、初列風切は手に対して鋭角をなしている。このような配列はミクロラプトルと非常によく似ている。いくつかの初列風切と次列風切は非対称な形にみえる。
このようにゼニュアンロンは前肢が短いわりに、ミクロラプトルとよく似た大きく複雑な構造の翼をもっている。この翼が何らかの航空力学的機能をもっていたかどうかは、生体力学的解析をしないとわからないが、体の大きさと前肢の短さを考えると可能性は小さいといっている。大型で前肢の短いゼニュアンロンは、小型で飛行性の祖先から進化し、祖先の翼の特徴を飛行のためではなく別の理由で保持しているかもしれない。大きく複雑な翼はディスプレイには有用だろうといっている。
系統解析の結果はちょっと意外なものであった。従来は、遼寧省のドロマエオサウルス類はミクロラプトル亜科Microraptorinaeとしてクレードをなすことが多かったが、ゼニュアンロンを加えて解析すると解像度が悪くなってしまった。今回の系統解析では、ゼニュアンロンとティアンユーラプトルが「前肢の短いドロマエオサウルス類」としてクレードをなすことはなかった。また遼寧省のドロマエオサウルス類が一つのクレードをなすこともなかった。代わりに、すべての遼寧省のドロマエオサウルス類と、ローラシアのドロマエオサウルス亜科やヴェロキラプトル亜科を含むクレードが大きなポリトミーをなした。この不確実性は広汎な収斂によるものである。現在のところ、遼寧省のドロマエオサウルス類が一つのクレードをなすのかどうか、その中に「前肢の短いドロマエオサウルス類」のグループがあるのかどうか、はっきりしないという。ミクロラプトル類が実は多系統になってしまうとすれば、ドロマエオサウルス類の系統関係に大きな影響がありそうである。
この論文には、化石の発見の経緯や地質の情報がないので、寄贈された標本だなと予想できる。学名はスン・ジェンユエン氏への献名で、「この美しい化石を研究のために保全した栄誉をたたえて」とあるので、てっきりこの人が寄贈者と思ったら、どうも違うようだ。博物館に寄贈したのは地元の農民で、素性を明かそうとしなかったとある。そうするとスン・ジェンユエン氏の役割は、農民が化石ブローカーに売ろうとしたのを思いとどまらせ、博物館に寄贈するように説得したということなのか。名前を出したくないということは、多分ああいうことなのだろうが、どんなドラマがあったのか、いろいろ勘ぐってしまう。
参考文献
Lu: , J. and Brusatte, S. L. A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution. Sci. Rep. 5, 11775; doi: 10.1038/srep11775 (2015).
コメント ( 2 ) | Trackback ( 0 )
デイノニクス(2)RPR モデル(猛禽獲物拘束モデル)続き
(前回の続きです。)
獲物を捕らえて動けないように固定したら、次は解体して摂食する作業である。その方法は現生の猛禽類では種類によって異なり、フクロウ類は獲物を丸呑みするが、タカ類とハヤブサ類は獲物を解体してから飲み込む。タカ類は獲物を両足の間に固定し、頭を下げてカギ状に曲がったクチバシで獲物の組織をつかみ、上方に引っ張って羽毛を抜いたり肉片を裂いたりする。著者らはデイノニコサウリアもタカ類と同様の姿勢で摂食したと考えた。
デイノニクス、ヴェロキラプトル、サウロルニトレステスの顎は特に頑丈ではなく、咬む力はそれほど強くないので、最初の攻撃や獲物を抑えるのには向いていない。RPRモデルでは、これらの顎はもっぱら解体作業に用いられたと考える。獲物は通常、かなり小さいもので、両足で完全に固定してあるので、顎にあまり負担はかからない。タカ類では咬む力が比較的弱いことが知られている。
獲物を解体する際の頭の方向や動きによって、ドロマエオサウルス類の特徴的な歯の形態が説明できるかもしれない。多くのドロマエオサウルス類では、後縁の鋸歯が前縁の鋸歯よりもずっと大きく、長くのびて先端がフック状に曲がっている。この形質は、派生的な白亜紀後期の種類(ヴェロキラプトルやサウロルニトレステス)で特に強まっている。RPRモデルでは、フック状の後縁の鋸歯は獲物を顎でグリップするのに効果的だったと考える。頭を両足の間に下げて獲物をくわえ、頭を上に引き上げると、獲物の組織を後縁の鋸歯に引っ掛けることになる。つまりこの鋸歯の形態は、獲物の組織を引きちぎる際のグリップを強めていた可能性がある。あるいは、獲物の羽毛や毛を引き抜くのに役立った可能性もあるという。
ドロマエオサウルス類の手の指はものを掴むのに適している一方、風切羽が生えていて動きが不自由にも見えることは依然として謎である。現生の猛禽類の行動を観察した結果、前肢の機能についてこれまで考えられていなかった仮説が浮かんできた。現生の猛禽類が足で獲物を抑えながら、翼で獲物を囲う行動:「マントリング」と呼ばれる姿勢がよく観察される。この行動は、獲物の逃亡を防いだり、他の捕食者に横取りされないように確保するためと考えられている。RPRモデルでは、前肢のカギ爪は、逃げようとする獲物を足元に戻すように掻き寄せるために用いられたと考える。これは前肢を足の近くに下げた時の手の角度とも一致する。
ドロマエオサウルス類は、大型の地上性のハンターでも「把握する足」を発達させたという話であった。この辺まで読んで、樹上性の小型の祖先が獲得した「枝をつかむための足」を地上に降りてからも活用した、という話だろうと思っていると、Fowler et al. (2011)はなんと、逆のプロセスを提唱している。
ミクロラプトルの研究では、足の多くの特徴、たとえば第 I 指が比較的遠位についている、足のカギ爪が強く湾曲している、遠位の末節骨以外の指骨が長い、などは樹上性への適応とされている。また基盤的な鳥類にみられる逆向きの第 I 指は、枝を握るための樹上性の適応と考えられている。Fowler et al. (2011)によると、これらのものを掴む、引っ掛ける機能に関わる特徴は、捕食のための適応でもある。ではどちらの役割が先なのかということである。Fowler et al. (2011)は、ものを掴むための適応は、小型の種類にも大型の種類(樹に登れない)にも同じようにみられることから、捕食性が先ではないかといっている。基盤的なデイノニコサウリアの中足骨が疾走に適していることも挙げている。そして樹上性の祖先が発達させた「ものを掴む足」を捕食性に転用したという可能性もあるが、そのためには地上性のハンターであるパラヴェス類の祖先が、樹上性の習性と適応を獲得し、その後地上に降りて失ったと考えなければならない。それよりも、「ものを掴む足」は地上性のハンターにおいて、もともと捕食のために進化し、のちに樹上性に転用された可能性が高いといっている。この辺りは微妙かなと筆者は思った。
さらに著者らは、パラヴェス類は捕食行動の間に「安定性はばたき」stability flapping を行ったと考えている。タカ類が大きめの獲物を捕獲して固定する際に、両足の第 II 指のカギ爪を獲物にしっかり食い込ませているので、体の安定を保つのに足を使うことができない。それを補うために、タカ類は盛んに「安定性はばたき」を行う。これはまず獲物の上に乗り、その位置を一定に保って、体重をかけて獲物を地面に固定するのに役立つ。微調整で済む場合は、翼を広げてときどき軽いはばたきを行うだけでよいという。
「安定性はばたき」は「はばたきが先モデル」 flapping first model を支持するという。これは、はばたき運動とそれによる上昇力の発生などは、本来飛行とは独立に進化できたというものである。アルカエオプテリクス、ミクロラプトル、シノルニトサウルスなどの基盤的パラヴェス類やデイノニコサウリアには、大きな翼がある。ヴェロキラプトルのような大きめの種類にも風切羽があったことが知られている。しかし飛翔の進化との関連では多くの議論がある。中途半端な翼が何の役に立つだろうか。「安定性はばたき」においては、比較的小さい気流や弱いはばたき能力でさえ役に立っている。アルカエオプテリクスや基盤的デイノニコサウリアのアスペクト比が小さい翼は、現生のタカ類の翼と似ている。タカ類は森林性の猛禽で急襲して獲物を捕らえ、頻繁に「安定性はばたき」を行う。短く幅の広い翼は操作性に優れており、「安定性はばたき」に適している。デイノニクスの前肢の動きは、鳥類のはばたきの形と似ているとされている。デイノニコサウリアが鳥と全く同じようなはばたき運動はできなかったとしても、「安定性はばたき」の際にそれに近い原始的なはばたきをすることは可能だったろう、といっている。タカ類には長い尾羽があり、「安定性はばたき」においてバランスをとるのに役立っている。デイノニコサウリアにも羽の生えた長い尾があり、同様にバランスをとるのに役立っただろうという。
Fowler et al. (2011) の論調は以上のような感じであるが、論文を読んでの私の感想は以下のようである。
似た形態をもつ現生動物の習性・行動との比較から入るというアプローチは面白く、成果も出している。ドロマエオサウルス類の足がタカ類と最も似ていること、獣脚類の足の形態が定量的に疾走型や把握型などのグループに分かれ、ドロマエオサウルス類は「把握する足」をもつことが認識された。デイノニコサウリアの中でトロオドン類とドロマエオサウルス類が別々の道をたどったことも示された。
テノントサウルスのような大型恐竜の背中に登ってカギ爪で腹を切り裂いた、というようなイメージよりは、ほとんどの場合自分よりずっと小さい獲物を捕食したというイメージの方が、はるかにリアルであり、獲物を捕獲した後はまさにRPRモデルのような姿勢で固定しただろうと納得できる。歯の特徴も、ナイフのように切断するというよりも少しずつ引きちぎる、引き裂くという動作を考えるとなるほどと納得した。バラウルは第 I 指のカギ爪も大きいが、RPRモデルのように固定するのには便利かもしれない。
しかし全体として、あまりにも現生の猛禽類とのアナロジーに頼りすぎているような気もする。そんなに何もかもタカ類と同じように考えなくてもよいのではないか。
ドロマエオサウルス類のカギ爪は、捕獲した獲物を固定するときにはRPRモデルのように役立っただろう。では獲物を捕らえるときには使わなかったのだろうか?走りながら小型の獲物を引っ掛け、押さえつけるのには十分役立ったのではないか。なぜ、最初に獲物を捕獲する段階の考察が抜けているのだろうか。先日、科博でオオタカやオウギワシの足の骨格を見たが、タカ類では第 I 指の方が大きめのようであった。また、「オオタカの狩り」の映像も見てきたが、タカ類にとっては飛行しながら獲物を掴んでかっさらう(スナッチ)がまず重要な動作のように思われた。この動き自体は、地上性の捕食者であるドロマエオサウルス類にはできない芸当である。そこで、著者らは獲物を捕らえた後の固定に集中してモデルを考えたのではないか。もっと最初に獲物を捕獲するところを考えてほしいと思った。
「安定性はばたき」も飛行できる動物であるタカ類がはばたきを行うのと、デイノニクスの体重で翼も不完全な前肢を動かすのとでは、勝手が違うのではなかろうか?そもそもデイノニクスくらいの大型種になると、小型の獲物を押さえつけるのに両足で獲物に乗っかっただろうか。よく描かれる肉食恐竜の捕食ポーズのように片足で押さえつけるので十分であれば、「安定性はばたき」は不要ではないか。小型の祖先の習性を引き継いでいるのかもしれないが。
まあいろいろと示唆に富むというか、さらなる研究の糸口になるような面白い研究に違いない。皆さんはこのモデルについてはどうお考えだろうか。
参考文献
Fowler DW, Freedman EA, Scannella JB, Kambic RE (2011) The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds. PLoS ONE 6(12): e28964. doi:10.1371/journal.pone.0028964
獲物を捕らえて動けないように固定したら、次は解体して摂食する作業である。その方法は現生の猛禽類では種類によって異なり、フクロウ類は獲物を丸呑みするが、タカ類とハヤブサ類は獲物を解体してから飲み込む。タカ類は獲物を両足の間に固定し、頭を下げてカギ状に曲がったクチバシで獲物の組織をつかみ、上方に引っ張って羽毛を抜いたり肉片を裂いたりする。著者らはデイノニコサウリアもタカ類と同様の姿勢で摂食したと考えた。
デイノニクス、ヴェロキラプトル、サウロルニトレステスの顎は特に頑丈ではなく、咬む力はそれほど強くないので、最初の攻撃や獲物を抑えるのには向いていない。RPRモデルでは、これらの顎はもっぱら解体作業に用いられたと考える。獲物は通常、かなり小さいもので、両足で完全に固定してあるので、顎にあまり負担はかからない。タカ類では咬む力が比較的弱いことが知られている。
獲物を解体する際の頭の方向や動きによって、ドロマエオサウルス類の特徴的な歯の形態が説明できるかもしれない。多くのドロマエオサウルス類では、後縁の鋸歯が前縁の鋸歯よりもずっと大きく、長くのびて先端がフック状に曲がっている。この形質は、派生的な白亜紀後期の種類(ヴェロキラプトルやサウロルニトレステス)で特に強まっている。RPRモデルでは、フック状の後縁の鋸歯は獲物を顎でグリップするのに効果的だったと考える。頭を両足の間に下げて獲物をくわえ、頭を上に引き上げると、獲物の組織を後縁の鋸歯に引っ掛けることになる。つまりこの鋸歯の形態は、獲物の組織を引きちぎる際のグリップを強めていた可能性がある。あるいは、獲物の羽毛や毛を引き抜くのに役立った可能性もあるという。
ドロマエオサウルス類の手の指はものを掴むのに適している一方、風切羽が生えていて動きが不自由にも見えることは依然として謎である。現生の猛禽類の行動を観察した結果、前肢の機能についてこれまで考えられていなかった仮説が浮かんできた。現生の猛禽類が足で獲物を抑えながら、翼で獲物を囲う行動:「マントリング」と呼ばれる姿勢がよく観察される。この行動は、獲物の逃亡を防いだり、他の捕食者に横取りされないように確保するためと考えられている。RPRモデルでは、前肢のカギ爪は、逃げようとする獲物を足元に戻すように掻き寄せるために用いられたと考える。これは前肢を足の近くに下げた時の手の角度とも一致する。
ドロマエオサウルス類は、大型の地上性のハンターでも「把握する足」を発達させたという話であった。この辺まで読んで、樹上性の小型の祖先が獲得した「枝をつかむための足」を地上に降りてからも活用した、という話だろうと思っていると、Fowler et al. (2011)はなんと、逆のプロセスを提唱している。
ミクロラプトルの研究では、足の多くの特徴、たとえば第 I 指が比較的遠位についている、足のカギ爪が強く湾曲している、遠位の末節骨以外の指骨が長い、などは樹上性への適応とされている。また基盤的な鳥類にみられる逆向きの第 I 指は、枝を握るための樹上性の適応と考えられている。Fowler et al. (2011)によると、これらのものを掴む、引っ掛ける機能に関わる特徴は、捕食のための適応でもある。ではどちらの役割が先なのかということである。Fowler et al. (2011)は、ものを掴むための適応は、小型の種類にも大型の種類(樹に登れない)にも同じようにみられることから、捕食性が先ではないかといっている。基盤的なデイノニコサウリアの中足骨が疾走に適していることも挙げている。そして樹上性の祖先が発達させた「ものを掴む足」を捕食性に転用したという可能性もあるが、そのためには地上性のハンターであるパラヴェス類の祖先が、樹上性の習性と適応を獲得し、その後地上に降りて失ったと考えなければならない。それよりも、「ものを掴む足」は地上性のハンターにおいて、もともと捕食のために進化し、のちに樹上性に転用された可能性が高いといっている。この辺りは微妙かなと筆者は思った。
さらに著者らは、パラヴェス類は捕食行動の間に「安定性はばたき」stability flapping を行ったと考えている。タカ類が大きめの獲物を捕獲して固定する際に、両足の第 II 指のカギ爪を獲物にしっかり食い込ませているので、体の安定を保つのに足を使うことができない。それを補うために、タカ類は盛んに「安定性はばたき」を行う。これはまず獲物の上に乗り、その位置を一定に保って、体重をかけて獲物を地面に固定するのに役立つ。微調整で済む場合は、翼を広げてときどき軽いはばたきを行うだけでよいという。
「安定性はばたき」は「はばたきが先モデル」 flapping first model を支持するという。これは、はばたき運動とそれによる上昇力の発生などは、本来飛行とは独立に進化できたというものである。アルカエオプテリクス、ミクロラプトル、シノルニトサウルスなどの基盤的パラヴェス類やデイノニコサウリアには、大きな翼がある。ヴェロキラプトルのような大きめの種類にも風切羽があったことが知られている。しかし飛翔の進化との関連では多くの議論がある。中途半端な翼が何の役に立つだろうか。「安定性はばたき」においては、比較的小さい気流や弱いはばたき能力でさえ役に立っている。アルカエオプテリクスや基盤的デイノニコサウリアのアスペクト比が小さい翼は、現生のタカ類の翼と似ている。タカ類は森林性の猛禽で急襲して獲物を捕らえ、頻繁に「安定性はばたき」を行う。短く幅の広い翼は操作性に優れており、「安定性はばたき」に適している。デイノニクスの前肢の動きは、鳥類のはばたきの形と似ているとされている。デイノニコサウリアが鳥と全く同じようなはばたき運動はできなかったとしても、「安定性はばたき」の際にそれに近い原始的なはばたきをすることは可能だったろう、といっている。タカ類には長い尾羽があり、「安定性はばたき」においてバランスをとるのに役立っている。デイノニコサウリアにも羽の生えた長い尾があり、同様にバランスをとるのに役立っただろうという。
Fowler et al. (2011) の論調は以上のような感じであるが、論文を読んでの私の感想は以下のようである。
似た形態をもつ現生動物の習性・行動との比較から入るというアプローチは面白く、成果も出している。ドロマエオサウルス類の足がタカ類と最も似ていること、獣脚類の足の形態が定量的に疾走型や把握型などのグループに分かれ、ドロマエオサウルス類は「把握する足」をもつことが認識された。デイノニコサウリアの中でトロオドン類とドロマエオサウルス類が別々の道をたどったことも示された。
テノントサウルスのような大型恐竜の背中に登ってカギ爪で腹を切り裂いた、というようなイメージよりは、ほとんどの場合自分よりずっと小さい獲物を捕食したというイメージの方が、はるかにリアルであり、獲物を捕獲した後はまさにRPRモデルのような姿勢で固定しただろうと納得できる。歯の特徴も、ナイフのように切断するというよりも少しずつ引きちぎる、引き裂くという動作を考えるとなるほどと納得した。バラウルは第 I 指のカギ爪も大きいが、RPRモデルのように固定するのには便利かもしれない。
しかし全体として、あまりにも現生の猛禽類とのアナロジーに頼りすぎているような気もする。そんなに何もかもタカ類と同じように考えなくてもよいのではないか。
ドロマエオサウルス類のカギ爪は、捕獲した獲物を固定するときにはRPRモデルのように役立っただろう。では獲物を捕らえるときには使わなかったのだろうか?走りながら小型の獲物を引っ掛け、押さえつけるのには十分役立ったのではないか。なぜ、最初に獲物を捕獲する段階の考察が抜けているのだろうか。先日、科博でオオタカやオウギワシの足の骨格を見たが、タカ類では第 I 指の方が大きめのようであった。また、「オオタカの狩り」の映像も見てきたが、タカ類にとっては飛行しながら獲物を掴んでかっさらう(スナッチ)がまず重要な動作のように思われた。この動き自体は、地上性の捕食者であるドロマエオサウルス類にはできない芸当である。そこで、著者らは獲物を捕らえた後の固定に集中してモデルを考えたのではないか。もっと最初に獲物を捕獲するところを考えてほしいと思った。
「安定性はばたき」も飛行できる動物であるタカ類がはばたきを行うのと、デイノニクスの体重で翼も不完全な前肢を動かすのとでは、勝手が違うのではなかろうか?そもそもデイノニクスくらいの大型種になると、小型の獲物を押さえつけるのに両足で獲物に乗っかっただろうか。よく描かれる肉食恐竜の捕食ポーズのように片足で押さえつけるので十分であれば、「安定性はばたき」は不要ではないか。小型の祖先の習性を引き継いでいるのかもしれないが。
まあいろいろと示唆に富むというか、さらなる研究の糸口になるような面白い研究に違いない。皆さんはこのモデルについてはどうお考えだろうか。
参考文献
Fowler DW, Freedman EA, Scannella JB, Kambic RE (2011) The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds. PLoS ONE 6(12): e28964. doi:10.1371/journal.pone.0028964
コメント ( 2 ) | Trackback ( 0 )
デイノニクス(2)RPR モデル(猛禽獲物拘束モデル)

Copyright Fowler et al. (2011)
デイノニクスくらい研究の歴史が長いと、当然ながら山のように文献があり、わりと重要と思われるものだけで10個くらいはある。全部はとても読めないので、興味深いと思える研究を探してみた。これは少し前の話題であるが、私は論文を読んでいなかったので、真面目に読んでみたら予想以上に面白かった。「デイノニクスの捕食生態学」というキャッチーな表題で、かなり説得力があるが突っ込みどころもあるような気がする、面白い仮説である。少し長いが論旨に沿って紹介してみたい。
デイノニクスといえば、恐竜のイメージを一新させた昔の「恐竜ルネッサンス」の象徴的な恐竜である。デイノニクスの群れがテノントサウルスを襲い、その背中に駆け上り、後肢のカギ爪で獲物の脇腹を切り裂いている・・・そのようなイメージが強い印象を与えてきた。デイノニクスのようなドロマエオサウルス類の後肢の大きな鎌状のカギ爪は、獲物の体を切り裂く、または駆け上るための適応であり、ドロマエオサウルス類は自分よりもはるかに大きな獲物を活発に襲撃し、殺害することに特化した捕食者であると想定されてきた。しかしこうしたデイノニクスの生態についての考えの多くは、推測に基づいている。発達した後肢の第 II 指のカギ爪は研究者の興味を引いてきたが、恐竜の爪の形態を、生態のわかっている現生動物と比較した研究はほとんどなかった。Fowler et al. (2011) はドロマエオサウルス類のカギ爪を含めた足の形態を、現生の猛禽類と詳細に比較することで、新しい仮説を提唱している。
Fowler et al. (2011) はまず、現生の猛禽類において、足の形態が捕食行動とどのように関連しているかを徹底的に解析した。その結果、タカ科Accipitridaeの猛禽類も顕著に大きな第 II 指のカギ爪をもっており、それは獲物の動きを封じること prey immobilisation に用いられていることがわかった。捕食者にとって、捕まえた獲物が逃げたり反撃したりしないように制圧することは大変重要である。現生の猛禽類ではそのための戦略はさまざまで、獲物の大きさによって変わってくる。小型の獲物の場合、足でつかむことで保定され、足の指で締め付けたりクチバシでつついたりする。小型の獲物を専門とするフクロウ類は、足の指で強く締め付けることに最も適応している。ハヤブサ類は獲物を動かなくするためにクチバシで脊髄をつついたり、頭を割ったりする。一方、足の中に収まらないような大きい獲物は、締め付けることはできない。大きな獲物の逃亡を防ぐため、猛禽は自分の体重をかけて獲物を地面に固定し、羽毛や毛を抜き始める。このときタカ類はカギ爪を用いる。タカ類は足の第 I 指と第 II 指に大きく発達したカギ爪をもっており、これで必死にもがく獲物を強く固定し、生きたまま捕食を始める。獲物は出血多量などによって絶命する。
Fowler et al. (2011) はこのような足の形態的特徴と捕食行動との関係を、絶滅した獣脚類にも応用することを考えた。現生の猛禽類のデータと比較することでデイノニコサウリアの足の機能形態を解析し、捕食行動と関連づけた。結論として、デイノニコサウリアの大きな第 II 指のカギ爪は、タカ科の猛禽と同じように獲物を固定するために用いられたと考え、RPR (Raptor Prey Restraint 猛禽獲物拘束) モデルとして提唱した。
まず種々の現生鳥類(タカ科、ハヤブサ科、コンドル科、フクロウ目、スズメ目など)で各指の末節骨の長さ、曲率、指骨の長さなどを測定したデータに、デイノニクスのデータを入れて比較すると、デイノニクスの足の形態はタカ科の猛禽と最も似ているという結果が得られた。これには大きな第 II 指のカギ爪や、その他の指骨の相対的な比率が寄与している。

また26種類の獣脚類について、各指の中足骨の長さ、末節骨の長さ、曲率、その他の指骨の長さなどを測定したデータを多変量解析で処理し、2次元のグラフを描くと、分類群や生活様式ごとにいくつかのグループに分かれた。走行性のオルニトミムス類とあまり走行性でないドロマエオサウルス類は大きく離れてプロットされた。トロオドン類はドロマエオサウルス類よりもむしろオルニトミムス類に近い位置にきており、より走行性を示している。中間的な位置にはティラノサウルス類やアロサウルス類が位置しており、アルカエオプテリクスもドロマエオサウルス類より中間的な位置にきた。
デイノニコサウリアの第 II 指のカギ爪が大きく強く湾曲していることは、現生のタカ類と同様に獲物の固定に用いられることを示唆している。足の内側にあり比較的短い指についているので、第 II 指のカギ爪は力を加えるのに最も役立つ。現生の肉食の鳥(カラスやハゲワシであっても)では第 II 指のカギ爪は食物を固定するのに用いられている。活発に捕食するタカ類やハヤブサ類は、屍肉食の種類に比べて強く湾曲した第 II 指のカギ爪をもっている。多くの肉食の獣脚類でも第 II 指のカギ爪が最も大きく、同じように固定するのに用いられたと思われる。一方、肉食でない現生の鳥類では第 III 指のカギ爪が最も大きく、肉食のものほどカーブしていない。同様に、二次的に植物食となったオルニトミムス類やアヴィミムスでは第 III 指のカギ爪が最も大きく、すべてのカギ爪で曲率は非常に小さい。このように第 II 指のカギ爪の湾曲と相対的な大きさは、肉食性、あるいは捕食性の指標となりうる。

足(中足骨と指骨)の相対的なプロポーションは、走行性やものを掴む(把握)などの機能に応じて変化する。エミューのような現生の走鳥類や、オルニトミムス類のように走行に適応した獣脚類では、第 III 指が太く、遠位の指骨が短くなり、側方の指( II と IV )が短く同じくらいの長さである。デイノニコサウリアと基盤的アヴィアラエの足では逆の傾向がみられる。第 IV 指が長くなり、遠位の末節骨以外の指骨が長くなり、第 II 指は過伸展している。これらの形質は、走行よりも把握に適している。
中足骨が長いことは、歩幅が大きくなり走行には適しているが、足の指でものを掴む力は弱くなる。フクロウ類は中足骨が短いことで、握る力が強くなっている。基盤的なパラヴェス類(基盤的トロオドン類シノヴェナトル、基盤的ドロマエオサウルス類シノルニトサウルス)は、比較的長い中足骨をもつ。これはもともと走行性だったことを示す。派生的なトロオドン類では完全にアルクトメタターサルな中足骨となり、さらに走行に適応している。それに対して、デイノニクス、サウロルニトレステス、ヴェロキラプトルのような派生的なドロマエオサウルス類は、長い中足骨を失って代わりに短く太い中足骨を進化させた。このことは、原始的な走行性の中足骨が、トロオドン類ではさらに走行性に適応していったのに対して、ドロマエオサウルス類では(走行性を犠牲にしてでも)強く掴む方向へと進化したことを示している。

指骨と指骨の間の関節面の形状も、足の使い方についての戦略と関係している。蝶番関節ginglymoid articulation は関節の動きを一方向に限定するので、ねじれに対する抵抗が強い。非蝶番関節 non-ginglymoid articulation (ローラー関節 roller joint) はねじれに対して抵抗が少なく柔軟性を示し、走行性の種類の指によくみられる。走行性の平胸類やオルニトミムス類では、主に体重を支える第 III 指の指骨に非蝶番関節 がある。現生の猛禽類では、すべての指骨間関節が蝶番関節であり、獲物の動きに抵抗して強く握ることに関係している。派生的なドロマエオサウルス類では、すべての中足骨と指骨間の関節が蝶番関節である(第 IV 中足骨を除く)。一方派生的なトロオドン類であるトロオドンでは、蝶番関節は一部の関節に限られる。第 III 指の指骨には非蝶番関節があり、第 IV 指には弱い蝶番関節があるのみである。このことからもトロオドン類がドロマエオサウルス類よりも走行性に適していることがわかる。
デイノニクスの足で関節の可動範囲を調べてみると、最大限に曲げなくても、足で「こぶし」を握ることができることがわかった。現生鳥類では第 I 指が第 III 指と対向するが、デイノニクスでは内側を向いた第 I 指が第 IV 指と対向し、第 II 指と第 III 指は平行に動く。これはフクロウが第 IV 指を移動した形と似ているという。他のドロマエオサウルス類の足もデイノニクスの足と似ていることから、「把握する足」はドロマエオサウルス類全体に共通する形質と考えられる。
(長いので一旦切ります。つづく)
コメント ( 0 ) | Trackback ( 0 )
デイノニクス(1)頭骨の特徴

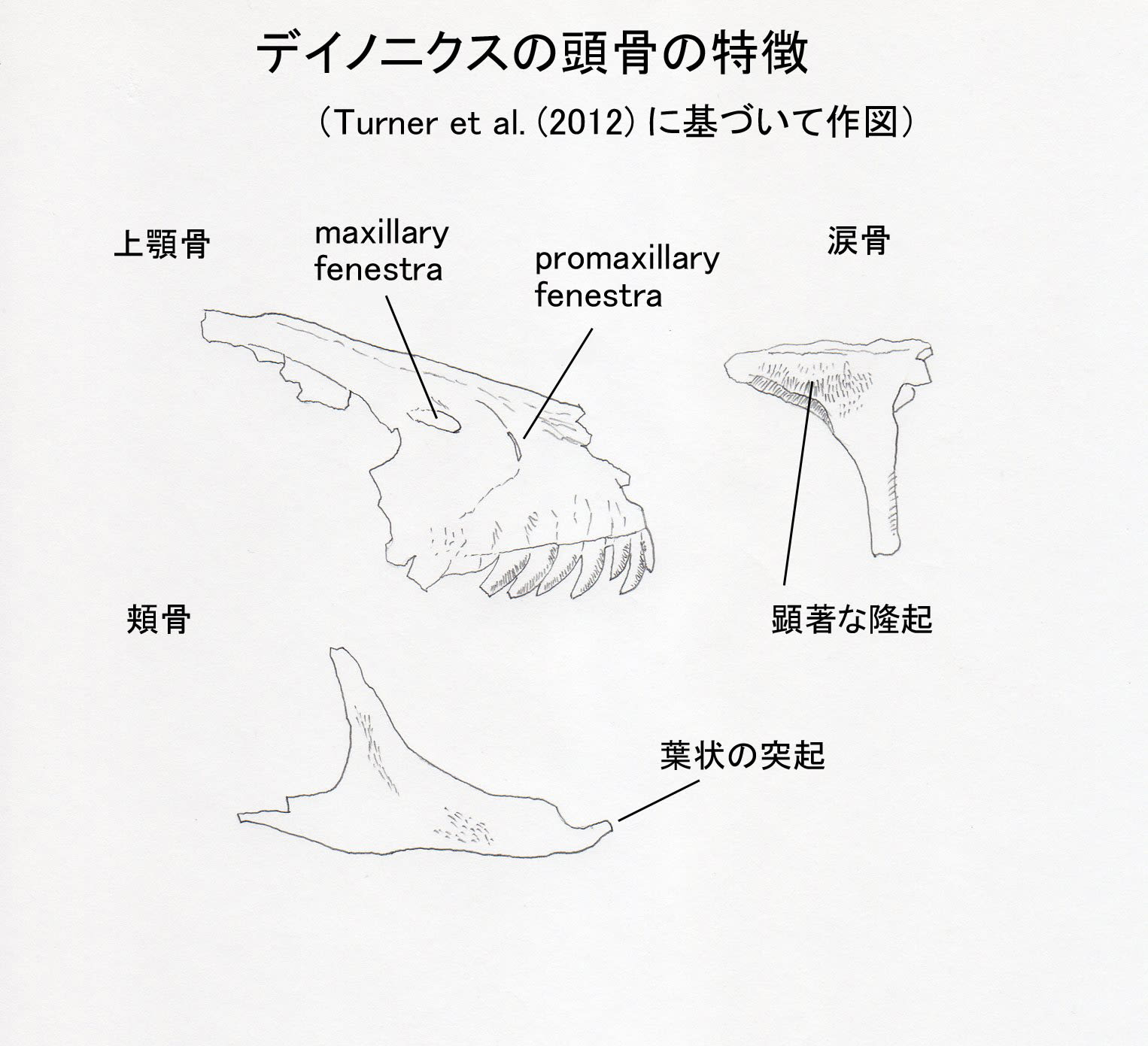
大きい画像 (図1)
Turner et al. (2012) によると、デイノニクスは以下の形質の組み合わせと固有形質により識別される大型のドロマエオサウルス類である。15本の上顎骨歯、4本の非対称な亜切歯形の前上顎骨歯、16本のほとんど同形の歯骨歯、すべての歯に前縁と後縁の鋸歯がある、すべての歯において後縁の鋸歯が前縁の鋸歯のほとんど2倍の大きさである、頬骨の前方突起の前端が葉状lobateである、涙骨に顕著な隆起bossがある、前前頭骨が非常に縮小している、maxillary fenestra がpromaxillary fenestra よりも背方にある、大きく幅広い踵骨。
ここで、上顎骨の前眼窩窩にある2つの孔、maxillary fenestra とpromaxillary fenestraの位置関係に注目してみよう(図1)。デイノニクスではmaxillary fenestra がpromaxillary fenestra のレベルよりも完全に上方にあり、細長い楕円形をしている。これはOstrom (1969)の図でも確かにそうなっている。Turner et al. (2012) の写真は、Ostrom (1969) の記載した上顎骨YPM5232そのものであるから当然である。また、他の研究者の論文で、いろいろなドロマエオサウルス類の上顎骨を並べて孔のパターンを比較した図があり、確かにデイノニクスの孔の配置は他の種類と識別できる特徴的なものであることがわかる。つまり、デイノニクスの重要な特徴の一つである。

図2 ちーたんの館のデイノニクス
そこで、丹波のちーたんの館にあるデイノニクス全身骨格の頭骨を観察してみた。(図2)
……あれ?
これは一体、どういうことだろう。maxillary fenestra が妙に丸く、promaxillary fenestraと同じ高さにあるように見える。少なくとも、不正確ではなかろうか。このキャストは世界中の博物館に納品されている商品のはずである。それなのにデイノニクスの重要な特徴が。。。
アリオラムスに続いて、またパンドラの箱を開けてしまったのだろうか。
参考文献
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
Ostrom, J.H. (1969). Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Bulletin of the Peabody Museum of Natural History, Yale University, New Haven, CT 30, 1-165.
(追記)
最初に掲げた「頭骨の特徴」の線画は、Turner et al. (2012) の写真をみて描いたものなので、微妙な凹凸までは表現されていない。Ostrom (1969) の記述には、maxillary fenestra (second antiorbital fenestra) は半円形とあるので、先の線画よりはもう少し広がっているかもしれない。しかし、maxillary fenestraはpromaxillary fenestra より明らかに上(背方)にある。これは間違いない。

科博のデイノニクス
そこで年明け早々に、科博にデイノニクス詣でに行って観察してきた。その結果‥‥
maxillary fenestraは半円形で、promaxillary fenestra より明らかに背方にある。つまり、この点について科博のデイノニクスは正確である。このキャストは前肢の向きが哺乳類的になっていておそらく古いものと思われるが、上顎骨の特徴は正確に作られているようだ。タッチパネルの解説中の頭骨図はOstrom (1969) そのものなので、もちろん正確である。
ちなみに、数年前に東海大学自然史博物館に行ってデイノニクス全身骨格を見た。そのとき頭骨のアップは撮らなかったが、科博のデイノニクスと同じもののように見える。
コメント ( 0 ) | Trackback ( 0 )
ダコタラプトル
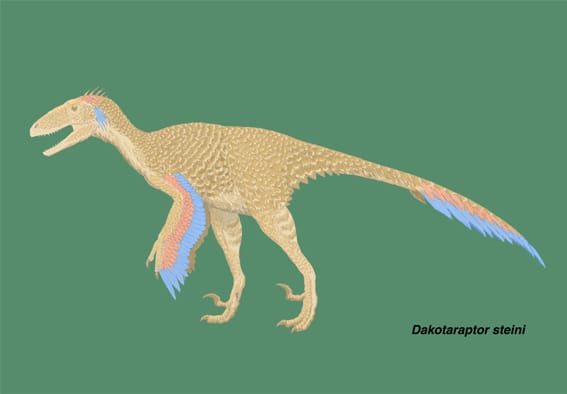

大きい画像
ダコタラプトルは、白亜紀後期マーストリヒト期(ヘル・クリーク層Hell Creek Formation)に米国サウスダコタ州に生息した大型のドロマエオサウルス類で、2015年に記載された。ほとんどが小型ないし中型のドロマエオサウルス類の中で、例外的に大型化した種類の一つであり、北アメリカではデイノニクス、ユタラプトルに続いて3番目に発見された大型ドロマエオサウルス類となった。また世界的に見ても最も生息年代の新しいドロマエオサウルス類であるという。
ホロタイプPBMNH.P.10.113.Tは、主に四肢の骨を含む成体の部分骨格で、頭骨はない。見つかっているのは1個の胴椎の椎体、10個の尾椎、叉骨、左右の上腕骨、左右の撓骨、左右の尺骨、右の中手骨IとII、3つの断片的な左の指骨、右の大腿骨、左右の脛骨、左の距骨と踵骨、左の中足骨II-IV、右の中足骨IV、右足の第II指と第III指の末節骨である。
参照標本として、より華奢な個体の右の脛骨、左の距骨と踵骨、叉骨、分離した歯などがある。
ダコタラプトルの特徴としては、1)例外的に大型のドロマエオサウルス類で、推定全長5.5 mに達する、2)足の第III指の末節骨の屈筋結節flexor tubercleが縮小している、3)足の第III指の末節骨の外側溝が、遠位側半分にわたって完全に閉じて骨質のチューブになっている、4)足の第II指と第III指の末節骨の腹側に、鋭いキールがある、5)fibular crestが長く、細く、その高さが長さの9%以下である、6)fibular crestの近位端がフック状である、などがある。(注:2)と3)については、論文のDiagnosisには第IV指と書いてあるが、誤植か直し忘れと思われる。本文を読むと第III指であるとわかる。Wikipedia英語版に第III指とあるのは正しい。)
左の尺骨は長さ36 cmで、よく保存されている。(尺骨+上腕骨)/大腿骨の比率は1.22で、ドロマエオサウルスやデイノニクスと近く、バンビラプトルほど大きくない。尺骨は長く、断面が円形で、他の多くのドロマエオサウルス類(ユタラプトルとアキロバトルを除いて)よりもがっしりしている。近位の関節面は破損しているが、それ以外の部分はデイノニクスやドロマエオサウルスと形態学的に似ている。尺骨は自然にカーブしているが、これはより直線的なバンビラプトルとは異なりデイノニクスと似ている。
尺骨の縁には10個の楕円形の隆起が並んでいる。これらの等間隔に並んだ突起は、羽軸こぶquill knobs(あるいは尺骨乳頭ulnar papilli)と考えられる。これらの間隔から、あと5個の羽軸こぶがあったと考えられ、ダコタラプトルの尺骨には全部で15本の風切羽があったと思われる。これはアルカエオプテリクスの12本、ヴェロキラプトルの14本、ミクロラプトルの18本と対応する。
ダコタラプトルの後肢のプロポーションは、同じくらい大型のユタラプトルのがっしりした後肢とは似ておらず、より小さいデイノニクスと似ている。つまり大型にもかかわらず、下肢が細長く疾走に適している。
脛骨は大きいが細長い骨で、ずっと太いユタラプトルの脛骨とは似ておらず、より小型のドロマエオサウルス類の脛骨と似ている。脛骨の長さは67.3 cmで、これまでに知られているドロマエオサウルス類の中で最も長い。ダコタラプトルの脛骨/大腿骨の比率は1.21で、ユタラプトルの1.00よりも大きくデイノニクスの1.10 に近い。(ドロマエオサウルスは1.37、バンビラプトルは1.39である。)
足の第II指の末節骨は完全に保存されており、ドロマエオサウルス類に特徴的な、典型的な鎌状のカギ爪である。この末節骨は、腹側の関節面から先端まで直線的に測ると16 cm、背側の曲がりに沿って測ると24 cmの長さである。この末節骨は大腿骨の長さの約29%あり、デイノニクスの23%よりも大きい。ダコタラプトルのカギ爪は、がっしりしていてユタラプトルと似ており、強く湾曲したデイノニクスのカギ爪とは異なっている。屈筋結節はユタラプトルと同様にがっしりとして顕著であり、関節面の腹側から下に張り出している。
足の第III指の末節骨もほとんど完全に保存されている。直線的に測ると7 cm、曲がりに沿って測ると9 cmである。末節骨の遠位部の断面は涙形teardrop-shapedで、腹側に鋭いキールがある。屈筋結節はほとんどないくらいに縮小している。外側溝は、遠位側で閉じて骨質のチューブになっている。この第III指の末節骨は、他のドロマエオサウルス類のものとは形がかなり異なるが、病理的な特徴ということはないだろうか。1個体の右側しかないのが残念である。せめて左右そろっていればもう少し確実なのだろう。
ダコタラプトルの歯は、強く側扁し、中程度ないし強く後方にカーブし、鋭く尖り、後縁の鋸歯が大きいという、一般的なドロマエオサウルス類の歯の形態をしている。ダコタラプトルの歯を他のドロマエオサウルス類の歯と定量形態学的に比較してみると、歯の形のプロポーションは他のドロマエオサウルス類と同様であったが、絶対的なサイズは群を抜いて大きいことがわかった。またダコタラプトルの鋸歯の密度は、ドロマエオサウルスとデイノニクスに最も近いことがわかった。歯の大きさ(歯冠の高さ)に対して鋸歯の密度をプロットしたグラフを描くと、ダコタラプトルはユニークな位置にくる。ダコタラプトルの歯冠の高さはティラノサウルスの集団の下端と重なるが、鋸歯の密度はドロマエオサウルスやデイノニクスと同様に大きく、ティラノサウルスとは全く重ならない。ダコタラプトルの値は他のどの獣脚類とも重ならないので、確実に見分けることができるという。
著者らの系統解析の結果では、ダコタラプトルはドロマエオサウルスと最も近縁で、次いでユタラプトル、アキロバトルと近縁な位置にきた。デイノニクスはさらに外側にきている。
これまでヘル・クリーク層の肉食恐竜といえば、大型のティラノサウルス類と小型のマニラプトル類に2分され、中間的なサイズの捕食者は知られていなかった。かなり大型で翼をもつダコタラプトルの発見によって、ヘル・クリーク層の動物相に対する我々のイメージは大きく変わったとしている。ダコタラプトルはおそらく、小型のマニラプトル類の獲物よりは大きく、大型のティラノサウルス類の獲物よりは小さい動物を捕食したと考えられる。ドロマエオサウルス類が群れで狩りをした可能性も考えると、ダコタラプトルが実際に小型のティラノサウルス類、つまりティラノサウルスの亜成体やナノティランヌスと獲物をめぐって競合する場面もあったかもしれない、と述べている。
ドロマエオサウルス類であるダコタラプトルが羽毛をもっていたことは十分予想されることであるが、このような大型のドロマエオサウルス類の尺骨に羽軸こぶが見つかったのは初めてであり、風切羽からなる翼をもっていたことがわかる。これはヴェロキラプトルでいわれているように、飛行のための翼を発達させた祖先の系統から受け継いだものである可能性が最も高いとしている。ダコタラプトルの行動・生態上の翼の役割としては、抱卵、求愛行動、なわばり行動などが考えられる。
Wikipedia等にも詳しく書かれているが、これはラプトル好きの人にはたまらない、エキサイティングな発見でしょう。大型で翼をもち、後肢が長く、カギ爪も大きいということで、白亜紀末の生態系を描くCG映像などでは活躍しそうである。
参考文献
DePalma, Robert A., Burnham, David A., Martin, Larry D., Larson, Peter L., Bakker, Robert T. (2015). The First Giant Raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. Paleontological Contributions (14).
コメント ( 0 ) | Trackback ( 0 )
ヴェロキラプトル


大きい画像
このサイトはドロマエオサウルス類が手薄だったので、勉強しようと思っていたところに、前述の総説が出てくれて大変助かった。これは200ページもあるので全部読むのは大変骨が折れるが、現時点でのドロマエオサウルス類の系統研究の状況がまとめてあり、ドロマエオサウルス類の好きな人には必見の資料である。
ところで皆さんは、ヴェロキラプトルの特徴をご存知だろうか。私は知らなかったので、調べた。もちろん、後肢の第2指のカギ爪、恥骨が後方を向く、尾椎が固定されて動かない等はドロマエオサウルス類の特徴であってヴェロキラプトルの特徴ではない。
Turner et al. (2012) では、ヴェロキラプトルの頭骨についてはBarsbold and Osmo´ lska (1999)、胴体についてはNorell and Makovicky (1997, 1999)の形質をほぼ採用している。
ヴェロキラプトルは、以下の形質の組み合わせと固有形質を示す中型のドロマエオサウルス類であるという。
上側頭窩(と上側頭窓)が亜円形で、側方に湾曲したsupratemporal arcadeで縁取られている。前頭骨が長く、眼窩の位置で幅に対して長さがほとんど4倍くらいあり、また頭頂骨のほぼ4倍の長さがある。前眼窩窓の前縁が広く丸い。maxillary fenestra が後方に開いた凹みの中にない。前上顎骨の上顎骨突起が長く、外鼻孔の後縁を越えて伸びている。歯骨の丈が非常に低く、その丈は長さの1/8から1/7である。歯骨の腹側縁が凸形にカーブしている。第1、第2前上顎歯が第3、第4より大きい。脳函の側壁に深いprootic recessがある。さらに叉骨、恥骨、座骨にそれぞれ固有形質があるようである。
頭骨の写真の説明には、さらに別の形質も記されている。maxillary fenestraが前眼窩窩の前縁から離れている(これは明らかにツァーガンとの区別のためだろう)、上側頭窩に余分の凹みがある、である。
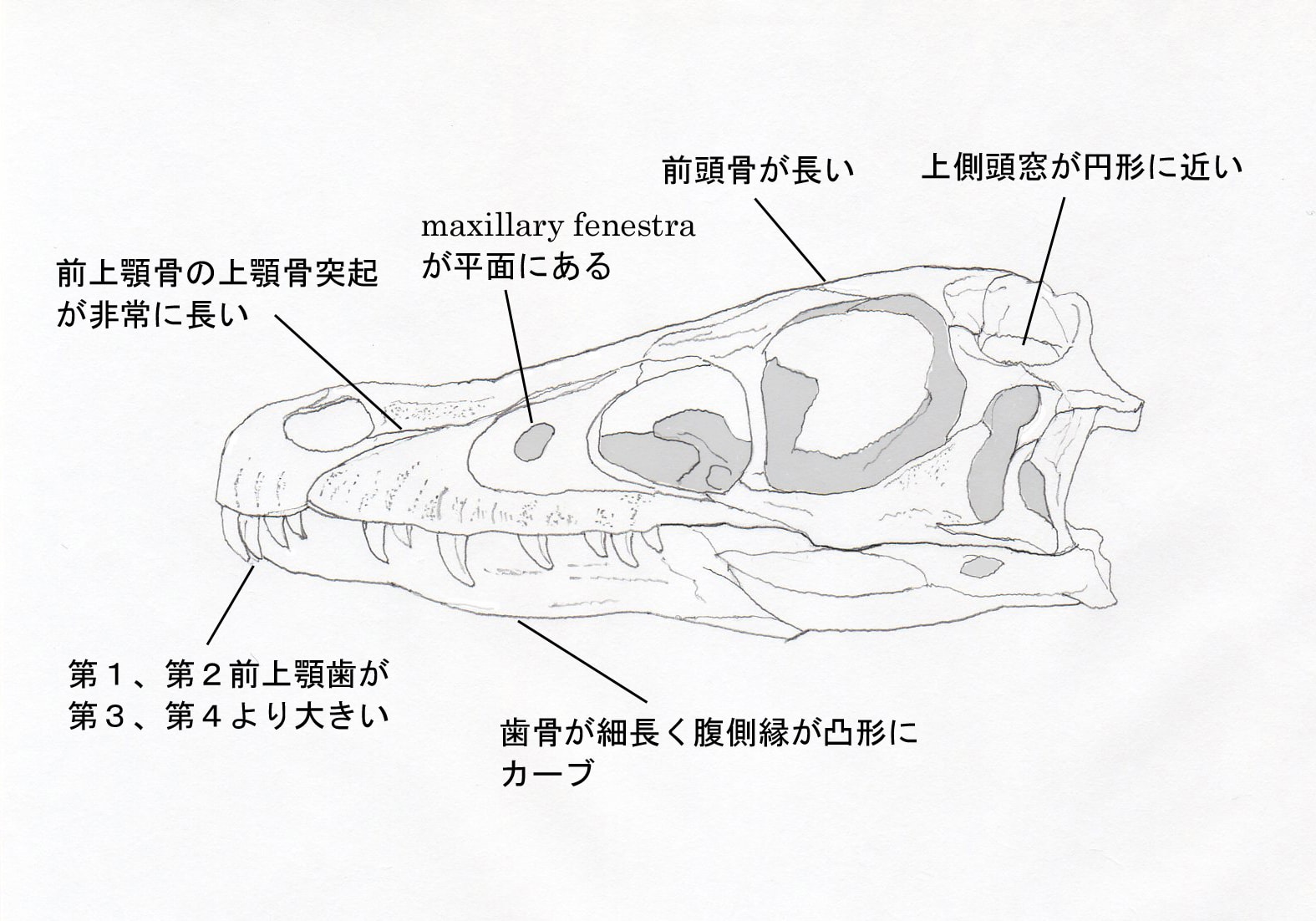
ドロマエオサウルス類は30種類も報告されているとはいっても、完全に近い頭骨が保存されているのはヴェロキラプトルやツァーガンを含めて数えるほどしかない。対応する骨が見つかっていない種類とは比較できないわけで、特徴といっても、どの種類と比較していっているのかが問題だろう。
Barsbold and Osmo´ lska (1999)によると、上側頭窩(と上側頭窓)が円形に近いというのはデイノニクスとの違いを述べているのであり、デイノニクスでは上側頭窩は細長く、まっすぐなsupratemporal arcadeで縁取られているという。前頭骨が長く、長さが幅の4倍というのは、デイノニクス、サウロルニトレステス、ドロマエオサウルスと比較している。サウロルニトレステスでは長さが幅のせいぜい3倍であり、ドロマエオサウルスでは2倍である。デイノニクスでは長さが幅の3倍で、頭頂骨の2倍であるという。
前上顎骨の上顎骨突起は、デイノニクスでは外鼻孔の後端を越えていないという。これについては、(Turner et al. (2012)にも特に書かれていないが)ツァーガンやリンヘラプトルではヴェロキラプトルと同様にみえる。
復元イラストや模型の作成にも関わることであるが、Turner et al. (2012)は一つ、Barsbold and Osmo´ lska (1999)とは異なる点について言及している。多くの研究者が、ヴェロキラプトルの鼻骨は中程が凹んでいる、つまり鼻先が「上を向いている」感じの顔と考えているが、これはアーティファクトであるという。鼻骨の前端部分は中央部よりも幅が広いので、ヴェロキラプトルの模式標本のように側面から押しつぶされた場合には、鼻先が上を向いて見えるのである、といっている。実際Turner et al. (2012)の復元頭骨図では、ヴェロキラプトルもツァーガンも、吻が直線的に描かれている。しかし、彼らのヴェロキラプトル頭骨はどうも細長すぎるように見える(ツァーガンよりも明らかに細長く描かれている)。鼻骨をまっすぐにするあまり、眼窩の上つまり額のあたりで急角度になっているようにみえるが、本当にこうなのだろうか。
Turner et al. (2012) はホロタイプだけでなく、格闘化石の頭骨も側面からつぶれていると考えている。しかしホロタイプは明らかにつぶれることで吻が上を向いているが、格闘化石の頭骨は多少つぶれているとしても、かなり保存が良いようにみえるのであるが。Barsbold and Osmo´ lska (1999)の復元頭骨図や、それを参考にしたとみられるVAP社やフェバリットの頭骨モデルも、鼻先はそれほど極端に上向きというわけではない。額から前方に向かってゆるやかに下りていき、水平な鼻先に移行するのでゆるく屈曲しているが、この方が自然な感じもする。
参考文献
Barsbold, R. & Osmdlska, H. 1999. The skull of Velociraptor (Theropoda) from the Late Cretaceous of Mongolia. Acta Palaeontologica Polonica 44, 2, 189-219.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ | 次ページ » |