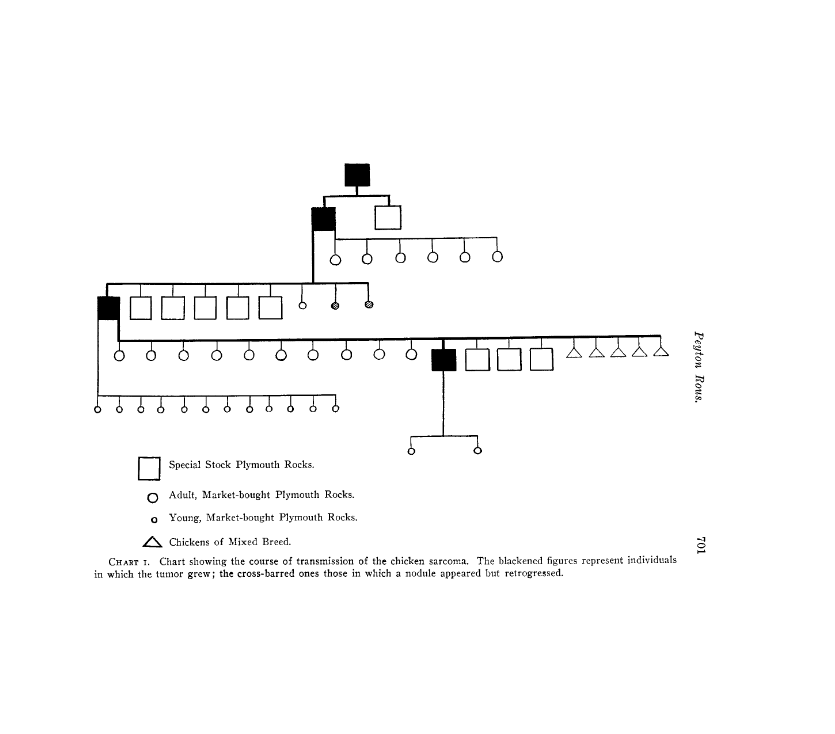
林房雄著『四つの文字』より (「白夫人の妖術」昭和38年11刷 文庫判 に収録)
彼は南京政府の部長ー日本流にいえば大臣であった。
略
当時、私は旅行者として北京に滞在していた。太平洋戦争の第二年目で、日本軍の占領地区は概して平和であったが、私は私なりに、日本の将来、アジアの運命について憂いをもっていた。
略
「ここでしたよ。」大臣はくりかえした。「殺しましたよ。二百人・・・・二百五十人。みんな若い学生でしたな。共産党員だということでしたが、その中の何人が共産党員であったか、わかりませんな。機関銃でなぎ倒したのですよ。・・・殺されたのは、私ですよ。ふふ、ふふふふ。」
引用終わり
最近、中国人ユーチューバーのLeiが、戦時中に毛沢東が日本に軍隊派遣を要請したという手紙がある話をしていた。
上記の大臣は、国民政府軍である。南京大虐殺は、日本対中国ではなく、日本・国民政府軍対共産党なのか?毛沢東の要請があったなら、国民政府軍対日本・共産党ではないのか。