講義 救急・集中治療領域における免疫学的考察と臨床研究の重要性
名古屋大学大学院医学系研究科 救急・集中治療医学分野
松田直之
はじめに
免疫(immunity)は,異物に対する生体の防御機能であり,ローマ共和制時代の元老院議員の課税や罰則の法的免除を意味するimmunitasを語源とする1)。生体の免疫反応は,大きく2つに分類され,生まれながらに既に獲得している自然免疫(innate immunity)や天然免疫(native immunity),そして微生物や癌などの異物により刺激されて防御能力を獲得する適応免疫(adaptive immunity)の2つのパターンが知られている。
このような免疫は,白血球系細胞と上皮細胞などの協調と調和の中で行われる。自然免疫では,好中球,マクロファージ,ナチュラルキラー細胞(NK細胞)などの食細胞と補体が重要な役割を担う。これらの食細胞には,異物を認識するToll-like受容体やスカベンジャー受容体などの細胞内情報伝達系が存在する。一方,適応免疫は,Bリンパ球とBリンパ球が産生する抗体が中心的役割を担う。抗体は,ウイルスや毒素などの異物の暴露から2〜4日遅れて,Bリンパ球より産生される。
虚血,細胞死,感染,異物などの生体侵襲は,交感神経と副交感神経の自律神経バランス,炎症性サイトカインと抗炎症性サイトカインのサイトカインバランス,ホルモン分泌変化において,免疫担当細胞に細胞数および機能変化をもたらす。外傷,crash症候群,環境異常症の管理においては虚血と壊死の側面から,感染症学については異物に対する観点から,ミトコンドリア細胞死とともに生体侵襲を捉え,基礎研究では創薬基盤形成を目標とし,臨床研究の側面からはこれらの管理学を発展させる必要がある。本稿は,生体侵襲の与える免疫機構への影響を総論としてまとめ,適時,内容をアップグレードさせることで,救急・集中治療医がその専門性をより一層に高めるために,生体侵襲と免疫の関係についての理解を深めることを目的とする。
1.自然免疫と細胞性免疫
生体が展開する免疫反応は,発生面では自然免疫と適応免疫,機能面では細胞性免疫と体液性免疫の大きく2つに分類される。これらに関与する免疫担当細胞は,骨髄の造血幹細胞(hematopoietic stem cell:HSC)2)に由来し,HSCの分化したものである(図1)。細胞性免疫は,主に好中球,マクロファージ,樹状細胞が担当する。液性免疫は,Bリンパ球の抗体産生が重要であり,ウイルスなどの異物に対する中和と排除をもたらす。
細胞性免疫として貪食能力を持つ細胞は,食細胞(phagocyte)と呼ばれる。食細胞は,好中球,マクロファージ,樹状細胞である。好中球は,ヒト成人の末梢血で40~70%(約65%)を占め,ヒトの自然免疫における中心を担う。マクロファージは,単球が分化したものであり,組織に移行する中で,ミクログリア(脳),肺胞マクロファージ(肺),クッパー細胞(肝臓),メサンギウム細胞(腎臓),破骨細胞(骨),ランゲルハンス細胞(皮膚),類上皮細胞,巨細胞などとして機能する。また,樹状細胞は,末梢に存在する時期の未成熟な状態では微生物や異物を取り込む働きを持ち,末梢で勉強すると成熟型樹状細胞として組織からリンパ組織へ移動し,さらに長い樹状突起を伸し,二次リンパ組織内でTリンパ球と結合し,Tリンパ球を活性化させる。Tリンパ球を活性化させる際に,樹状細胞の役割は大きい。このような食細胞において,異物との関わりを外傷管理や手術管理で考えることは重要であり,食細胞機能がどのように修飾されるかの評価は診療において重要である。
組織における上皮系細胞,血管内皮細胞,そして,免疫担当細胞として好中球,マクロファージ,幼若型樹状細胞のような食細胞は,Toll-like受容体3-5)やスカベンジャー受容体6-8)を発現し,自然免疫や死細胞の除去に寄与している。スカベンジャー受容体は,異物や死細胞の貪食を行うものであり,炎症期における重要な役割を担う。スカベンジャー受容体6-8)は,表1にように直接に結合する自然免疫型受容体と補体や抗体によってオプソニン化された粒子に反応するオプソニン受容体に区分されている。
一方,好塩基球,好酸球,肥満細胞は,消化管,呼吸器,尿管の粘膜上皮における生体防御に関与している。これらの好塩基球,好酸球,肥満細胞は,直接に異物を認識できるが,貪食能はなく,細胞質内の顆粒分子を細胞外に放出することで,異物を除去する。寄生虫などの大きな異物に対して効力を持つ。
2.適応免疫と体液性免疫
適応免疫と体液性免疫に関与するリンパ球は,血液中では総白血球数の20~40%(約30%)であり,骨髄で成熟するBリンパ球と,胸腺で成熟するTリンパ球9-11)に分類される。リンパ球数が減少する病態は,低栄養と交感神経緊張であり,ウイルス感染症やエイズでも低下する。リンパ球のうち,液性免疫として抗体を産生するのはBリンパ球である。Tリンパ球は,細胞傷害性T細胞(cytotoxic T cell,キラーT細胞)とヘルパーT細胞(helper T cell)の2つに分類され,細胞傷害性T細胞は,ウイルスやレジオネラなどの細胞内寄生微生物を傷害させる。ヘルパーT細胞は,B細胞の抗原産生を活性化させる補助的な作用を持ち,さらに好塩基球,好酸球,肥満細胞の活性化に関与する。また,リンパ球の中には,細胞膜上に抗原特異的受容体を発現させていないナチュラルキラー細胞(NK細胞)12, 13)が存在する。ヘルパーT細胞やNK細胞は,異物を認識する際に,主要組織適合遺伝子複合体(major histocompatibility complex:MHC)を用いる特徴がある。
3. MHC分子の発現と機能
ヒトのMHCは,MHCクラスⅠ分子14, 15)とMHCクラスⅡ分子16 ,17)として2つのクラスターに分類されており,これらは遺伝子領域6p21.3で転写される。このゲノム領域は,免疫に関与する多くの分子を産生する領域であり,免疫応答の中心をなす領域である。翻訳後のMHCは,α鎖,β鎖および結合するペプチドの3量体であり,MHCクラスⅠ分子のβ鎖はβ2ミクログロブリンである。結合できるペプチド長は,MHCクラスⅠ分子でアミノ残基8~10個,MHCクラスⅡ分子で10~20個である。MHCはそれぞれ,細胞傷害性T細胞とヘルパーT細胞の機能において重要な役割を担う。
MHCクラスⅠ分子14, 15)は,細胞傷害性T細胞だけではなく,ほぼすべての細胞に存在しており,細胞質内でプロテアーゼにより消化されたベプチドと結合し,細胞膜上へ輸送する機能を持つ(図2)。一方,MHCクラスⅡ分子16 ,17)は,B細胞,マクロファージ,樹状細胞,NK細胞に高密度で発現しており,エンドソームで分解されて産生されたペプチドを細胞膜上に運搬し,ヘルパーT細胞による補助作用を強化する働きをする。例えば,B細胞では,抗体を細胞膜表面に膜型受容体として持つため,ここに結合した抗原をエンドサイトーシスとして細胞内へ運搬し,リソソームでペプチドに分解されるとMHCクラスⅡ分子により細胞膜上に運搬され,結合したヘルパーT細胞がB細胞の形質細胞への分化を誘導し,抗体産生を高める(図3)。また,マクロファージではMHCクラスⅡ分子を発現しており,マクロファージに内で消化されたペプチドをマクロファージの細胞膜上に発現させ,ヘルパーT細胞との連動によりマクロファージを活性化させる(図4)。このように,MHCクラスⅡ分子は,細胞内のペプチドを細胞膜上に運搬し,細胞膜上に発現させ,B細胞,マクロファージ,樹状細胞, NK細胞の機能を高める作用を担う。
また,MHCには,CD4やCD8などの共同受容体(co-receptor)が存在し,胸腺におけるT細胞の分化を補助している。CD8は,MHCクラスⅠ分子の共同受容体であり,細胞傷害性T細胞の異物に対する認識を高める。一方,CD4は,MHCクラスⅡ分子の共同受容体であり,マクロファージ,樹状細胞,およびB細胞の異物認識を高める。
4.補体の活性化と機能
補体18 ,19)は,自然免疫として細胞貪食に関与するほか,抗原のオプソニン化,標的細胞への細胞膜傷害性分子複合体(membrane attack complex)18-20)の活性化,抗体産生促進などを主作用とする。この補体系は,約30種類の血清蛋白と膜蛋白で構成されている。C1~C9の補体蛋白, B因子,D因子,マンノース結合レクチン関連セリンプロテアーゼMASP(-1, -2, -3),プロペルジンなどが代表的分子である(表2)。補体の活性化は,①抗原・抗体複合体による古典的経路,② 病原菌細胞膜に結合した抗体による第2経路,③ マンノース結合レクチンやリガンド結合型フィコリンなどによるレクチン経路の3つの経路(図5)が知られており,補体受容体(表3)を介した反応として説明できる。補体欠損による病態としては,感染症罹患率の増加が知られているほか,C1,C2, C4の欠損により古典的経路やレクチン経路が障害されると全身性エリテマトーデスの発症が高まることが知られている。
5. T細胞の活性化
T細胞は,胸腺内で成熟し,血液,二次リンパ組織,輸出リンパ管を循環しており,二次リンパ組織において成熟樹状細胞のMHC複合体と結合することによってエフェクターT細胞として活性化する。まず,胸腺においてT細胞は,Runx21)とTh-POX22)の2つの転写因子の関与により,CD8とCD4の発現制御が行われていることが確認されている。Runx21)は,T細胞においてCD8を発現させ,細胞傷害性T細胞としての分化をもたらす。一方,Th-POX22)は,T細胞にCD4を発現させ,ヘルパーT細胞としての分化をもたらす。このような過程において,成熟樹状細胞はナイーブT細胞に対してIL−2受容体α鎖(CD25)とIL-2の転写を高め,T細胞の成熟と活性化に関与する。T細胞におけるIL-2産生に関与する転写活性因子としては,① NFAT(nuclear factor of activated T cell),② NF−κB(nuclear factor-κB),③ AP-1(activator protein-1),④ Oct1の4つの関与が知られている。
このような状況において,CD4陽性ヘルパーT細胞は,Th1,Th17,Th2,Tregに分化する細胞内情報伝達シグナルを持つ(図6)。 Th1細胞は,IFN-γを産生し,マクロファージ,NK細胞,細胞傷害性T細胞の病原体排除機能を増強させる。Th17細胞は,IL-17を産生する細胞であり,IL-17RA/RC受容体の存在する好中球の活性化に関与する。一方,Th2細胞は,IL-4産生を特徴とし,好酸球,好塩基球,肥満細胞などの上皮系バリア機能を誘導する。Tregは,胸腺で誘導される内在性Treg(natural Treg:nTreg)と末梢で分化誘導される誘導性Treg(inducible Treg:iTregs)に分類されており, nTregは CD4+ CD25high であるT細胞として,FoxP3転写因子活性により誘導される。Tregは,IL-10およびTGF-βを産生することで細胞傷害性T細胞などの攻撃性を抑制するが,炎症回復期の線維芽細胞などの増殖にも関与している。
6.生体侵襲におけるDANPSとPAMPS
生体の細胞傷害や微生物との反応においては細胞性免疫が働くが,産生される炎症性サイトカインや炎症性分子により炎症という別な表現形が現れる。これらの反応は, damage-associated molecular patterns(DAMPs)と総称され,微生物との反応においてはpathogen-associated molecular patterns(PAMPs)と呼称され5, 23),このような2012年レベルのテクニカルタームの出現により,多くの言葉を説明に用いずに共通概念を描くことができるようになった。現在,2016年の段階では,虚血,細胞死,ミトコンドリア死などに伴い生体内で増加する内因性分子の受容体反応も解明されてきている。Toll-like受容体,nucleotide-binding oligomerization domain(NOD), NOD-like receptors (NLRs),AIM2-like receptors(ALRs),retinoic acid-inducible gene-Ⅰ(RIG-Ⅰ)like receptors(RLRs),C型レクチン受容体などは,主要なDAMPs受容体であり,細胞性免疫と炎症を考える上で重要である。 Toll-like受容体5)は,PAMPs受容体としてのみではなく,DAMPs受容体としての機能も持つ(表4)。
7.免疫能に対する生体侵襲の考察
生体侵襲については,私は2000年より,①虚血,②細胞死(壊死),③感染,④異物の4つとして,生体への侵襲の入力を説明してきた。結果としては,交感神経・副交感神経の活性バランス,サイトカイン産生,ホルモン分泌の表情(現れ方)の観察が重要である。外傷や手術などにおいては,直接的な組織傷害,虚血再灌流,組織乾燥などの影響により,DAMPs産生に伴うDAMPs受容体反応5, 23)として炎症が高まるばかりか,食細胞機能が変化する。外傷侵襲は,Th1/Th2バランスを低下させることが知られており24, 25),細胞性免疫低下として好中球やマクロファージなどの食細胞機能が低下し,DAMPsの組織内クリアランスの低下や,微生物や腫瘍細胞などに対する防御能が低下する可能性に注意する。さらに,DAMPsの蓄積は,組織・上皮系細胞の炎症性シグナルとして,急性肺傷害,急性腎傷害,播種性血管内凝固などの多臓器不全の誘引となる可能性を持つ5)。
このような状態の集中治療管理では,プロポフォール26, 27)やデクスメデトミジン28)などを鎮静剤として用いるが,これらはTh2有意に傾いたTh1/Th2バランスをTh1方向に改善させる傾向がある。鎮痛薬として使用されるモルヒネやフェンタニールは,オピオイドμ受容体を介してCD4陽性ヘルパーT細胞数を低下させ,CD8陽性細胞傷害性T細胞を増加させることが知られている29-31)。手術領域では,ASA physical status class 3あるいはclass 4の肺小細胞癌に対する24症例の前向き研究-32)として,硬膜外麻酔やフェンタニールによる術中・術後の疼痛管理で,術後1日におけるTh1/Th2バランスを術前と同等に維持できるという結論である。
生体侵襲期において交感神経活性を抑制することの意義は,血行動態の安定化や酸素需要の適正化に加えて,①コルチゾルの分泌抑制,②白血球系細胞の機能維持にある。生体侵襲による交感神経緊張は,代謝性アシドーシスの呼吸性代償,つまり過換気応答などの影響も受け,青斑核などからのノルエピネフリンの放出を介して視床下部からのCRH(corticotropin-releasing hormone)および下垂体前葉からのACTH(adrenocorticotropic hormone)の分泌を高め,副腎皮質からのコルチゾル分泌を高める。これらは,食細胞機能を低下させる可能性や多能性分化細胞の機能不全を導く可能性がある。血漿コルチゾルは,2~18μg/dLレベルの日内変動で維持されているが,生体侵襲により反応性にコルチゾルが高い状態では,免疫抑制状態となることを確認できる。フェンタニール33),バルビタールやプロポフォール34),ミダゾラム35)やエトミデート36)などは,下垂体前葉からのACTH放出や副腎皮質からのコルチゾル分泌を抑制する。アドレナリンα2作動薬デクスメデトミジンも,副腎皮質束状帯におけるコルチゾル産生を抑制する37)。
生体侵襲の緩和を目的とする「集中治療管理」などにおいては,このような静脈麻酔薬濃度と免疫細胞機能との関連に洞察が必要である。さらに,好中球やリンパ球にはアドレナリン受容体が存在し,高濃度カテコラミン暴露により,好中球やリンパ球は細胞死を起こす38, 39)。出血により虚血が生じる場合には,ミトコンドリア機能が変化し,あるいはミトコンドリア死により,DANPs反応として細胞性免疫が低下する。白血球除去の不確実な輸血によっては,輸血後10日でも細胞傷害性T細胞が減少することが報告されており40),輸血後に腫瘍免疫やウイルス免疫が低下する可能性についても洞察が必要である。
おわりに
生体侵襲における免疫管理は,早期退院および長期予後改善のために今後より一層に不可欠な急性期管理における学術となる。T細胞におけるT細胞受容体機能,B細胞の抗体産生機能,補体への作用,好中球などの食細胞機能,スカベンジャー受容体機能などを,自律神経,サイトカイン,およびホルモンの各バランスの観点より時系列で的確に評価していくことが必要である。急性期管理病態学を研鑽する中で,救急・集中治療の重要性として,免疫の研究を推進しなければならない。
文 献
1. A highly detailed compendium of developments in all aspects of allergy will be found in H. Schadewaldt, Geschichte der Allergic, 4 vols. Dustri-Verlag, Dusseldorf, 1979.
2. Cheng Y, Newell EW. Deep Profiling Human T Cell Heterogeneity by Mass Cytometry. Adv Immunol. 2016;131:101-34.
3. Joosten LA, Abdollahi-Roodsaz S, Dinarello CA, et al. Toll-like receptors and chronic inflammation in rheumatic diseases: new developments. Nat Rev Rheumatol. 2016;12:344-57.
4. Alvarado AG, Lathia JD. Taking a Toll on Self-Renewal: TLR-Mediated Innate Immune Signaling in Stem Cells. Trends Neurosci. 2016. 30005-4.
5. Matsuda N. Alert cell strategy in SIRS-induced vasculitis: sepsis and endothelial cells. J Intensive Care. 2016:23;4:21.
6. Penberthy KK, Ravichandran KS. Apoptotic cell recognition receptors and scavenger receptors. Immunol Rev. 2016;269:44-59.
7.Zani IA, Stephen SL, Mughal NA, et al. Scavenger receptor structure and function in health and disease. Cells. 2015;4:178-201.
8. Canton J, Neculai D, Grinstein S. Scavenger receptors in homeostasis and immunity. Nat Rev Immunol. 2013;13:621-34.
9. Jackson HJ, Rafiq S, Brentjens RJ. Driving CAR T-cells forward. Nat Rev Clin Oncol. 2016 ;13:370-83.
10. Ilyas S, Yang JC. Landscape of Tumor Antigens in T Cell Immunotherapy. J Immunol. 2015;195:5117-22.
10. Durinck K, Goossens S, Peirs S, et al. Novel biological insights in T-cell acute lymphoblastic leukemia. Exp Hematol. 2015;43:625-39.
11. Malissen B, Bongrand P. Early T cell activation: integrating biochemical, structural, and biophysical cues. Annu Rev Immunol. 2015;33:539-61.
12. Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. 2007;25:297-336.
13. Brennan PJ, Brigl M, Brenner MB. Invariant natural killer T cells: an innate activation scheme linked to diverse effector functions. Nat Rev Immunol. 2013;13:101-17.
14. Apcher S, Prado Martins R, et al. The source of MHC class I presented peptides and its implications. Curr Opin Immunol. 2016 ;40:117-22.
15. Schuren AB, Costa AI, Wiertz EJ. Recent advances in viral evasion of the MHC Class I processing pathway. Curr Opin Immunol. 2016 ;40:43-50.
16. Forsyth KS, Eisenlohr LC. Giving CD4+ T cells the slip: viral interference with MHC class II-restricted antigen processing and presentation. Curr Opin Immunol. 2016;40:123-9.
17. Montano-Frías JE, Vera-Massieu C, Álvarez-Martínez R, et al. MHC class II transcription is associated with inflammatory responses in a wild marine mammal. Infect Genet Evol. 2016;42:77-82.
18. Merle NS, Church SE, Fremeaux-Bacchi V, et al. Complement System Part I - Molecular Mechanisms of Activation and Regulation. Front Immunol. 2015;6:262.
19. Merle NS, Noe R, Halbwachs-Mecarelli L, et al. Complement System Part II: Role in Immunity. Front Immunol. 2015;6:257.
20. Morgan BP. The membrane attack complex as an inflammatory trigger. Immunobiology. 2016;221:747-51.
21. Collins A, Littman DR, Taniuchi I. RUNX proteins in transcription factor networks that regulate T-cell lineage choice. Nat Rev Immunol. 2009;9:106-15.
22. Setoguchi R, Tachibana M, Naoe Y, et al. Repression of the transcription factor Th-POK by Runx complexes in cytotoxic T cell development. Science. 2008;319:822-5.
23. Janeway CA Jr, Medzhitov R. Innate immune recognition. Annu Rev Immunol. 2002;20:197-216.
24. Decker D, Schondorf M, Bidlingmaier F, et al. Surgical stress induces a shift in the type-1/type-2 T-helper cell balance, suggesting down-regulation of cell-mediated and up-regulation of antibody-mediated immunity commensurate to the trauma. Surgery 1996;119:316-25.
25. Decker D, Springer W, Decker P, et al. Changes in TH1/TH2 immunity after endovascular and conventional infrarenal aortic aneurysm repair: its relevance for clinical practice. Eur J Vasc Endovasc Surg 2003;25:254-61.
26. Inada T, Yamanouchi Y, Jomura S, et al. Effect of propofol and isoflurane anaesthesia on the immune response to surgery. Anaesthesia 2004;954-9.
27. Helmy SA, Wahby MA, El-Nawaway M. The effect of anaesthesia and surgery on plasma cytokine production. Anaesthesia 1999;54:733-8.
28. Kim Y, Kang SH, Hong TH, et al. Effects of dexmedetomidine on the ratio of T helper 1 to T helper 2 cytokines in patients undergoing laparoscopic cholecystectomy. J Clin Anesth. 2014;26:281-5.
29. Mao M, Qian Y, Sun J. Morphine Suppresses T helper Lymphocyte Differentiation to Th1 Type Through PI3K/AKT Pathway. Inflammation. 2016;39:813-21.
30. Gao M, Sun J, Jin W, et al. Morphine, but not ketamine, decreases the ratio of Th1/Th2 in CD4-positive cells through T-bet and GATA3. Inflammation. 2012;35:1069-77.
31. Carr DJ, France CP. Immune alterations in chronic morphine treated rhesus monkeys. Adv Exp Med Biol. 1993;335:35–9.
32. Cata JP, Bauer M, Sokari T, et al. Effects of surgery, general anesthesia, and perioperative epidural analgesia on the immune function of patients with non-small cell lung cancer. J Clin Anesth. 2013;25:255-62.
33. Hall GM, Lacoumenta S, Hart GR, et al. Site of action of fentanyl in inhibiting the pituitary-adrenal response to surgery in man. Br J Anaesth. 1990;65:251-3.
34. Cohan P, Wang C, McArthur DL, et al. Acute secondary adrenal insufficiency after traumatic brain injury: a prospective study. Crit Care Med. 2005;33:2358-66.
35. Broadbear JH, Winger G, Woods JH. Self-administration of methohexital, midazolam and ethanol: effects on the pituitary-adrenal axis in rhesus monkeys. Psychopharmacology. 2005;178:83-91.
36. Absalom A, Pledger D, Kong A. Adrenocortical function in critically ill patients 24 h after a single dose of etomidate. Anaesthesia. 1999;54:861-7.
37. Maze M, Virtanen R, Daunt D, et al. Effects of dexmedetomidine, a novel imidazole sedative-anesthetic agent, on adrenal steroidogenesis: in vivo and in vitro studies. Anesth Analg. 1991;73:204-8.
38. Kohm AP, Sanders VM. Norepinephrine and beta 2-adrenergic receptor stimulation regulate CD4+ T and B lymphocyte function in vitro and in vivo. Pharmacol Rev. 2001;53:487-525.
39. Tan KS, Nackley AG, Satterfield K, et al. β2 adrenergic receptor activation stimulates pro-inflammatory cytokine production in macrophages via PKA- and NF-κB-independent mechanisms. Cell Signal. 2007; 19:251-60.
40. Innerhofer P, Luz G, Spötl L, et al. Immunologic changes after transfusion of autologous or allogeneic buffy coat-poor versus white cell-reduced blood to patients undergoing arthroplasty. I. Proliferative T-cell responses and the balance of helper and suppressor T cells. Transfusion. 1999;39:1089-96.
図と表の解説
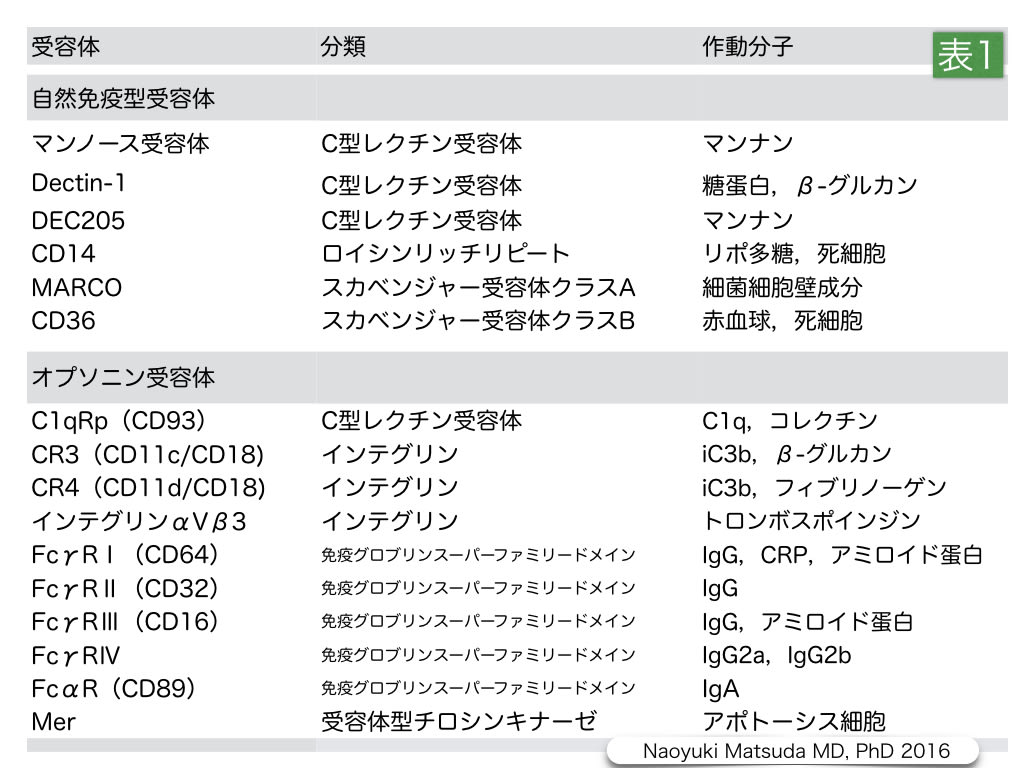
表1.スカベンジャー受容体の区分と作動分子
スカベンジャー受容体は,マンノース受容体やDectin-1のような然免疫型受容体と,CR3などのような補体やCD64のようなT細胞受容体などの抗体によって反応するオプソニン受容体に区分されている。

表2.補体の種類と機能

表3.補体受容体の種類と機能
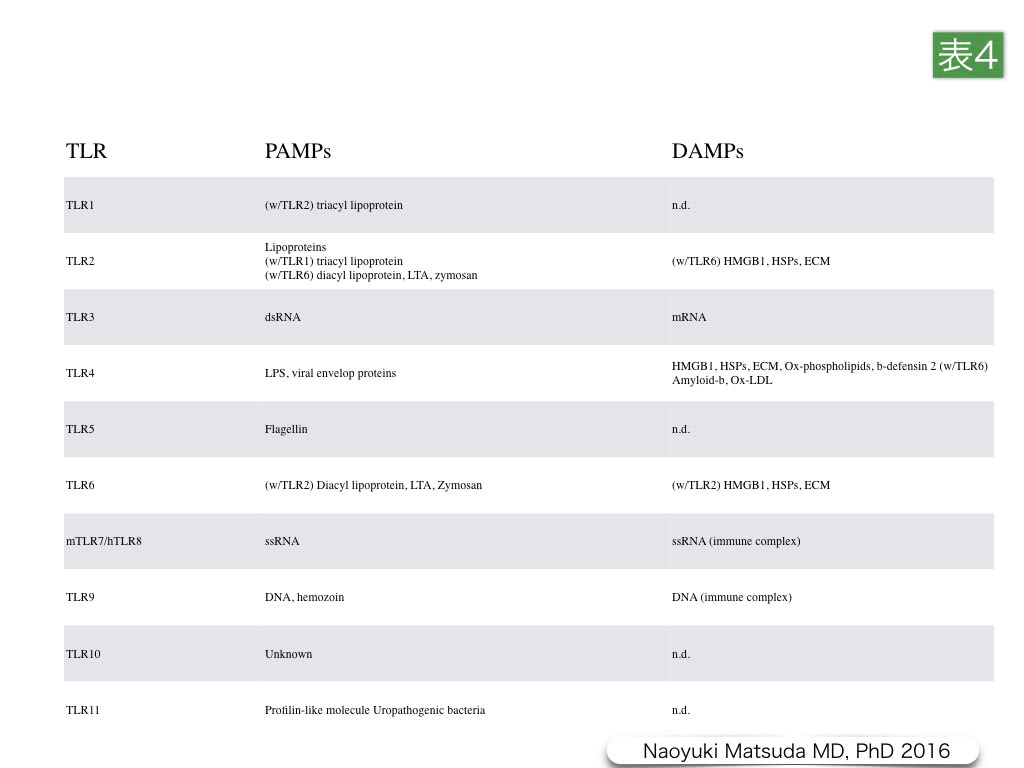
表4.Toll-like受容体におけるPAMPsとDAMPsリガンド
Toll-like受容体(TLR)は,単球,マクロファージ,樹状細胞,好中球,繊維芽細胞,内皮細胞,上皮細胞などのAlert細胞5)で炎症性分子の産生に関与する炎症性受容体であるとともに,免疫細胞で発現しているタイプI膜貫通型パターン認識受容体ファミリーの1つである。ヒトでは10種類のサブタイプ,マウスでは12種類のサブタイプが同定されている。TLRsは,細菌・カビ・原生生物・ウイルスなどの微生物の蛋白・核酸・脂質,炭水化物に保存されている病原関連分子パターン(pathogen-associated molecular patterns:PAMPs)を認識し,自然免疫や獲得免疫のトリガーとなるばかりではなく,damage-associated molecular patterns(DAMPs)として生体内分子の危機トリガーとして機能している。

図1. 造血細胞の分化
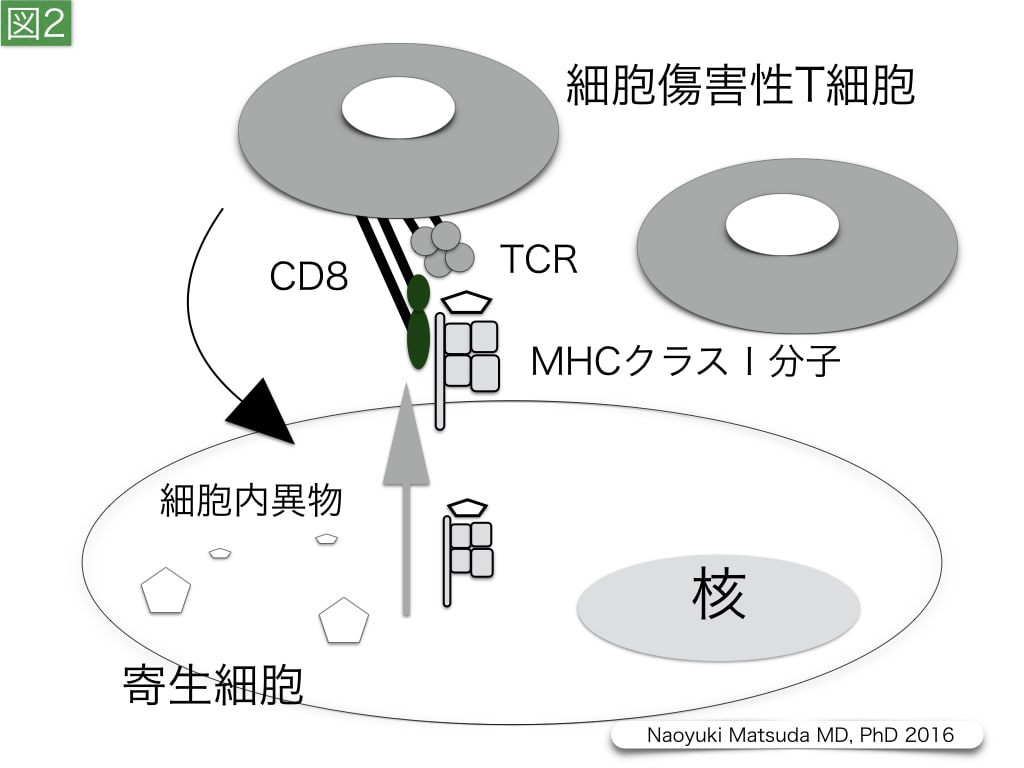
図2. 寄生細胞に対する細胞傷害性T細胞の作用
サイトメガロウイルスやレジオネラなどに感染された細胞では,CD8陽性の細胞傷害性T細胞の機能が期待される。寄生細胞では,寄生異物の断片分子や断片小胞がMHCクラスⅠ分子により細胞膜に運搬される。T細胞受容体(TCR)は,αおよびβのヘテロ2量体であり,CD3複合体と会合して,T細胞膜上に存在する。さらに,細胞傷害性T細胞では,TCRと細胞内異物・MHCクラスⅠ分子複合体との接着と認識に,CD8がco-receptorとして補助的に作用する。
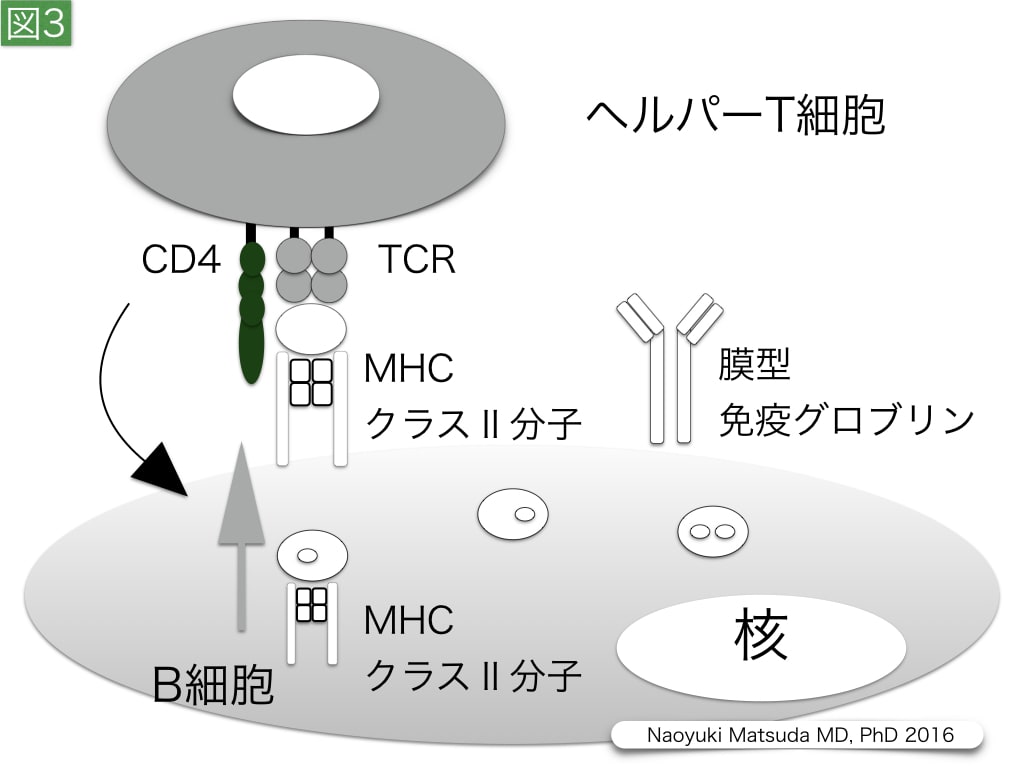
図3. B細胞に対するヘルパーT細胞の作用
B細胞は細胞膜上に抗体として膜型グログリンを発現することができ,膜型グロブリンを介して,結合した抗原を細胞内に取り込み,エンドソームでペプチドに断片化する。この抗原断片は,細胞質内でMHCクラスⅡ分子に補綴され,細胞膜上に運搬され,ヘルパーT細胞により認識される。ヘルパーT細胞には,T細胞受容体が存在し,CD4をco-receptorとして,B細胞の活性化を施し,この異物に対して特異的な抗体を産生する形質細胞やメモリー細胞(memory cell)への分化を促進させる。このようなB細胞分化において,抗体記憶に対する生体侵襲の影響の研究は不十分である。
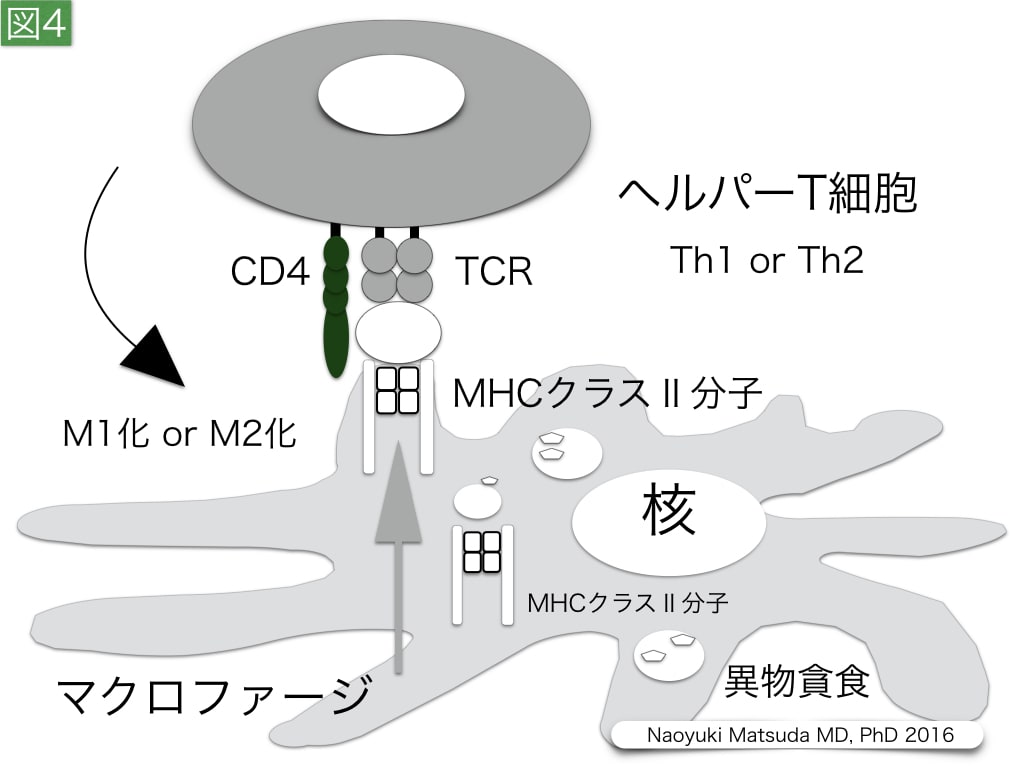
図4. マクロファージに対するヘルパーT細胞の作用
マクロファージ,好中球,未成熟樹状細胞は,食細胞として微生物などの異物を貪食する。主に組織に存在するマクロファージは,肺胞マクロファージ,クッパー細胞などのように,呼称を変えて呼ばれる。マクロファージは,貪食した分子をプロテアーゼでペプチドレベルに断片化する。この分解断片は,細胞質内でMHCクラスⅡ分子に補綴され,マクロファージの細胞膜上に運搬され,ヘルパーT細胞により認識される。ヘルパーT細胞のマクロファージの作用は,IFN-γを放出するTh1とIL-4およびIL-13の分泌を行うTh2細胞で異なる。Th1細胞は,IFN-γを介してマクロファージの貪食能と殺傷能をM1マクロファージとして高める。一方,Th2細胞は,IL-4とIL-13を介してM2マクロファージとしてマクロファージにIL-10やTGF-βなどの増殖性サイトカインの分泌をもたらす。M2マクロファージは,M1タイプの活性化マクロファージとは異なり,真菌認識を行うDectin-1,好酸球や好塩基球を誘導するCCL17,CCL18,CCL22などのケモカイン,そして線維芽細胞を増殖させるTGF-β,コラーゲン合成のために必要なプロリン産生を高めるアルギニン分解酵素アルギナーゼなどを産生する特徴があり,Th2有意のヘルパーT細胞バランスにより,炎症期に随伴した線維化期が遷延する危険性がある。

図5. 補体活性化の3つの経路
補体の活性化は,①抗原・抗体複合体による古典的経路,② 病原菌細胞膜に結合した抗体による第2経路,③ マンノース結合レクチンやリガンド結合型フィコリンなどによるレクチン経路の3つの経路が知られており,C3bによるC5b産生が重要な役割を担う。C3bはC3変換酵素により,C5と会合するとC5をC5aとC5bに分解できる。C5aは,C3aとともに,炎症性ペプチドとして作用する。一方,C5bは,C6,C7,C8細胞膜傷害性分子複合体(membrane attack complex)を形成し,さらにC9を細胞膜内に貫入させ,微生物膜に10 nmレベルの小孔を形成する。このような補体を介した小孔形成は,サイトメガロウイルス,コロナウイルス,ヘルペスウイルスのようなエンベロープを持つウイルス,ブドウ球菌などの厚いペプチドグリカン層を持つグラム陽性菌の除去に有効と考えられる。
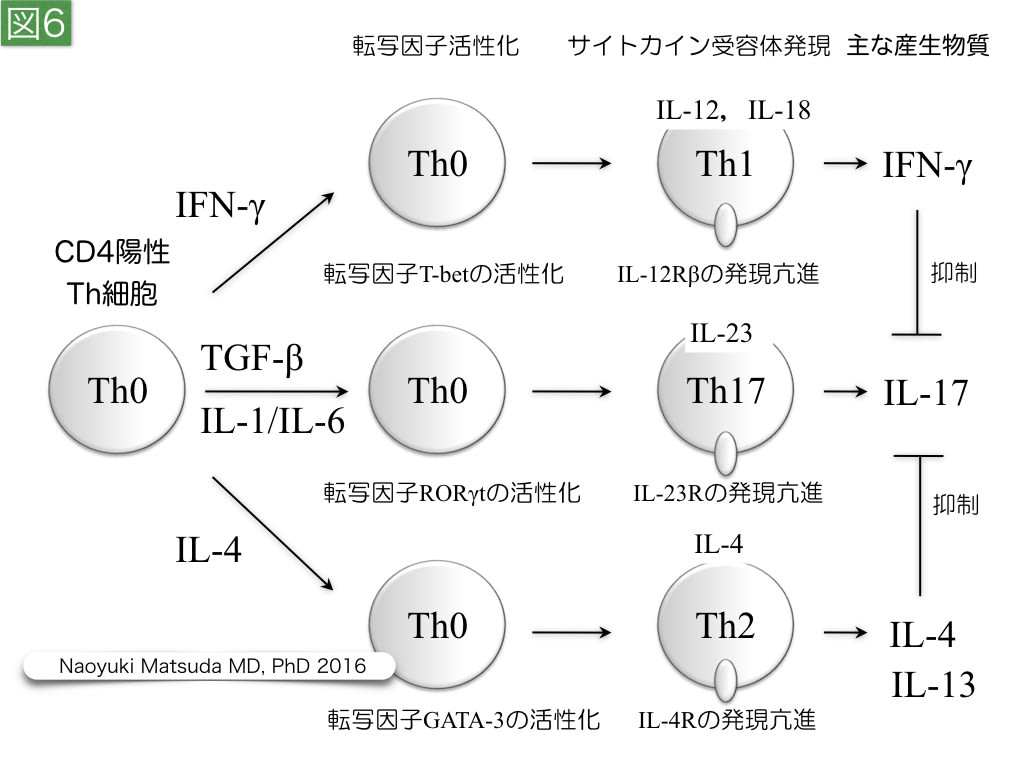
図6. ヘルパーT細胞の分化
ヘルパーT細胞は,T細胞受容体とCD4により,マクロファージ,B細胞,幼若型樹状細胞に結合し,これらの機能を修飾する。その作用は,分化形体により差異があり,転写因子活性とサイトカイン受容体シグナルにより,Th1,Th17,Th2などに分化し,産生し,放出するサイトカインを変える。