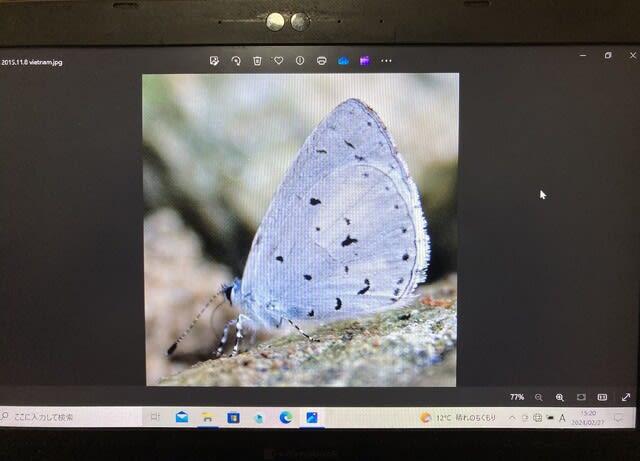
オニタビラコ(帰化植物)の花を訪れたオガサワラシジミ 小笠原母島乳房山山頂付近Apr.6, 1993
この連載というか一連の報文を書き始めた本来の目的は、昔(1970年代)にドッサリとっておいた、父島におけるオオバシマムラサキとシマムラサキへの産卵行動、および幼虫の周日活動などの写真&データを探し出して発表する事だった。が、それが出てこない。どこかにあるはずなんだが、ダンボールの中をいちいち探すのは至難の業である。ということもあって、まず御本尊ともいえるルリシジミをはじめとする関連種の紹介から初めて、それを行っているうちにオガサワラシジミ関連の資料も出てくるだろうと暢気に構えていたのだが、どだい端から探し出す努力をしていないものだから、出て来るわけがない。とりあえず今回はそれらの紹介は諦める。出てきたときに改め発表していきます。
ルリシジミの話に戻る。スギタニルリシジミやウラジロルリシジミ、あるいはアリサンルリシジミの一群も、オガサワラシジミ成立の何らかのカギを握ってはいるだろうけれど、やはり本命は御宗祖様のルリシジミであろう。見かけの差はともかく、様々な形質でオガサワラシジミに最も関連の深い種であることは確かだ。小笠原に最も近い地域、伊豆諸島南部や、南西諸島北部でも記録がある(分布の北上、ではなくて、南下ですね)。
ルリシジミは14亜種(Eliot&Kawazoeが選択したもの、記載自体はもっと沢山ある)が、ヨーロッパのほぼ全域から、北アフリカ、中東、中央アジア、ヒマラヤ、中国、台湾、日本海周辺、シベリア、北米、中米の、北半球冷温帯(いわゆる周北極圏)地域に広く分布し、唯一熱帯に相当するフィリッピン・ルソン島から亜種suguruiが記録されている。五十嵐邁氏がルソン島で採集し、Eliot&Kawazoeで新亜種記載が為された。
五十嵐氏の採集品には、もう一つの亜種が含まれている。北イラクのクルド地方で採集された、おそらく亜種hypoleucaに属する個体。その採集品は川副氏らには提供されていないのではないだろうか(標本、♂交尾器とも図示なし)。というのは、実は、それ以前に僕が譲り受けているのである。
経緯は忘れた(おそらく五十嵐氏が採集した当地のルリシジミの翅裏に青い鱗粉が顕著に覆うことを何かで知って氏に質問していたのだと思う)。氏の結婚式披露宴に出席した際、わざわざその標本を贈呈してくださったのだ。
早速♂交尾器をチェックしたのだが、ルリシジミとしてはごく平均的な形状、オガサワラシジミとの詳細な比較検討をしないままに今に至っている。川副氏がルリシジミを纏めると聞いた際、それを差し上げればよかったのだが、当然五十嵐氏から直接別個体が渡っているだろうと思っていた。考えてみれば貴重な資料だある、押し入れのダンボールのどこかに入っているはずなので、改めてチェックしたい。むろん、直接的には何の関係もないだろうけれど、オガサワラシジミ成立の何らかのヒントのようなものが暗示されているかも知れない。
小笠原が1968年に日本に返還された直後からの数年間、京都大学の小路義明氏たちによる詳細な調査が行われ、それと入れ替わるように僕が小笠原を訪れたのが1976年から1993年にかけてである(1976/1977/1979/1981/1988/1992/1993年)。写真やデータのかなりの部分を消失してしまったが、それでも今回運び込んだダンボール中に少なからぬ写真が残っているはずなのだが、現時点では見つけ出せないでいる。
1970年代後半には、各所に群がり飛んでいた父島のオガサワラシジミは、1980年代に入って激減、1988年の時点ではほとんど見ることが出来なくなってしまっていた。一方、以前から“大発生”という状況にはなかった母島では、余り顕著な変動はなく、少ないながらも確実に姿を見ることが出来た。
1988年の夏も小笠原に滞在していた。その年の春に最初の中国大陸行。ギフチョウ属やキマダラヒカゲ属の種をたっぷり撮影し、初夏、今後のフィールドを中国大陸に移そうと目論んで、大学に留学すべく東京の中国大使館を訪れたのだが、中卒はダメ!と情け容赦なく却下されてしまった(中卒どころか実質一年生までしか通っていなかったので、卒業証書も提出できなかった)。
それで諦めて、その夏も小笠原に渡っていたのである。8月末、役場に電報(まだ電話が充分に普及していなかった)。友子さんの父上からである。僕の代わりに大使館にウイスキーを2本携えて、再度申請に行った、すると許可が下りた由。新学期(向こうは9月)が始まるので、すぐに戻って来いと。それ以来、主戦場は中国に移ったのである。
中国と日本を行き来する間を縫って、1992年と1993年にも短期間母島を訪れた。92年には、北港道路(石門分岐点)路肩繁みに生えるタチアワユキセンダングサの花に多数吸蜜に訪れているのを撮影した。93年には乳房山山頂付近で、コオニタビラコの花に静止している個体を撮影(冒頭写真)。今思えば、70~80年代には、在来固有種シマザクラが主要吸蜜源(ほかにムラサキシキブ属各種)だったのが、90年代に入ってからは、帰化植物のタチアワユキセンダングサとコオニタビラコに代わってしまっていたのである。
70年代には余り記憶になかったタチアワユキセンダングサだが、80年代末から急激に増えだしたようである。ちなみに、当時並行してフィールドとしていた屋久島でも、80年代前半まではコシロノセンダングサばかりだったのが、90年代に入って久しぶりに訪れたら、まるで魔法のように、ほとんど全てがタチアワユキセンダングサに置き換わっていた。
1993年を最後に小笠原へは行っていない。「父島では絶滅したらしい」「母島でもほとんど見かけない」と言う声が聞こえてくる。
2018年、小笠原日本返還50周年記念ということで、インターネット・マガジン「現代ビジネス」にオガサワラシジミの話題について寄稿した。前後28回渡って掲載した「現代ビジネス」だが、蝶についての話題は後にも先にもこれ一回だけ。
国(東京都)や権威研究機関が“オガサワラシジミ”に対して行おうとしている“保護対策”に対する批判記事である(確かその数年前に朝日新聞社発行の科学雑誌“サイアス”にも同様のことを書いた)。国や都や自然保護機関が行おうとしている保全運動は安易に過ぎる。野生個体絶滅宣言、場所を移して飼育し、現地に再導入する試みなど、もってのほかだ、と。読者の反応は、28回の記事中、最悪(というか無関心)だったようである。
2020年になって、本土(多摩動物園、新宿御苑)での飼育系統個体も全滅、という情報。「それ見たことか」と言いたいところだが、そのような言い方は止めて置こう。看過するわけにはいかない、由々しき事態なのである。
オガサワラシジミの絶滅は、そん所そこらの“絶滅”とは訳が違う。
生物の種の絶滅は、その大半が次の2つのパターンに帰属する。
地域個体群の絶滅。分かりやすいのはトキの例だ。日本に於ける唯一の棲息地佐渡の個体群が絶滅した。しかし中国の秦嶺山地にも同じ種の個体群が健在、従って「種」が絶滅したわけではない(そのため人為的再導入が為された)。
蝶で言えば、オガサワラシジミと並んで、絶滅一番手と目されていた(オガサワラシジミと共に絶滅危惧第一類)日本本土の山地草原に棲む4種、オオルリシジミ、オオウラギンヒョウモン、ヒョウモンモドキ、ウスイロヒョウモンモドキは、それぞれ非常に限られた地域に、絶滅寸前の状況下で生き延び続けている。行政や自然保護団体が必死になって保護政策に取り組んでいる。これらの種は、ひと昔前までは、今よりもずっと広い範囲に分布していた。この数十年の間に急速に衰退して行ったのはオガサワラシジミと共通する。異なるのは、分布範囲が圧倒的に広い事。種としては、日本海を取り巻く極東アジアに広域分布しているのである。そのうちの南辺、すなわち日本列島に於いて急速に衰退しているのだが、北辺のロシア沿海部、朝鮮半島、中国東北地方などでは、必ずしも滅亡の危機に晒されているわけではない。日本国内では絶滅寸前だが、種としては健在なのである。

オオルリシジミ Glaucopsyche(Shijimiaeoides)divinus
長野県上田市 Jun.17,1990
現在では絶滅してしまった可能性のある産地。オオルリシジミはルリシジミと名が付くが、ルリシジミの仲間ではなく、カバイロシジミやゴマシジミのグループ。ちなみにオガサワラシジミの他に後翅裏基半部に顕著な青緑色鱗粉を備えるのは、日本ではこのオオルリシジミと、北海道やシベリアなどに分布するカバイロシジミぐらいである。

オオルリシジミ Glaucopsyche(Shijimiaeoides)divinus
長野県上田市 Jun.17,1990

オオルリシジミ Glaucopsyche(Shijimiaeoides)divinus
熊本県阿蘇山 May 21,1993
この産地は保護政策が取られていることから今も健在と思う。

オオルリシジミ Glaucopsyche(Shijimiaeoides)divinus
熊本県阿蘇山May 21,1993

オオウラギンヒョウモン Argynnis(Fabriciana)nerippe
大分熊本県境九重高原 Aug.9,1992

オオウラギンヒョウモン Argynnis(Fabriciana)nerippe
大分熊本県境九重高原Aug.9,1992

ヒョウモンモドキ Melitaea scotosia
広島県芸北町 Jul.10,1992
この産地における現在の状況は未詳(絶滅?)。

ヒョウモンモドキ Melitaea scotosia
広島県芸北町 Jul.10,1992

ヒョウモンモドキ Melitaea scotosia
広島県芸北町 Jul.9,1992

ウスイロヒョウモンモドキ Melitaea regama
島根県三瓶山 Jul.13,1993

ウスイロヒョウモンモドキ Melitaea regama
島根県三瓶山Jul.13,1993
その他にも、日本の各地で風前の灯状態にある、チャマダラセセリ、ウラナミジャノメ、ヤマキチョウなど絶滅危惧1B類の種も、中国に行けば大都市周辺で、地域によっては市街地の真っただ中で、ごく普通に見られたりする。なんで日本だけ衰退しているのだろうかという思いはあるにせよ、種としての絶滅とは意味が違う。
もう一つの“固有種絶滅”パターンは、確かに、離島や特殊環境に於いて“固有”であることには違いないにしろ、分類群(いわゆる“種”とはなっていても、実質的には品種程度)としては、ごく短い期間に成り立った変異集団に基づくもので、消滅の速度も速い可能性がある(いわゆる“絶滅種”の多くもそれに準じる)。極論を言えば、環境の推移次第で新たに再出現することもあり得るかもしれない集団。
オガサワラシジミの場合は、その両パターンとは明確に異なる。正真正銘“種”の絶滅である。問題の大きさが桁違い、例えて言えば、マンモスの絶滅が、今目の前で為されているようなものである。
ルリシジミやスギタニルリシジミが共通祖先種であることには間違いないが、♂交尾器の比較に基づけば、明らかに独立した固有分類群だ(例えて言えば、絶滅マンモスとアフリカゾウやインドゾウとの関係)。そのことを鑑みれば、公的機関の声明や報道は、余りに淡泊かつ無責任で、違和感を覚える。
通常、新たに種が形成されるのには、数百万年の時間が必要とされる。数百万年間、小笠原という空間は存在したのか。普通に考えれば、こんな小さな陸塊が水没せずに存在し続けることは有り得ないような気もする。ただ、ハワイの生物相の由来(詳細は別途に説明予定)が、単に洋島として捉えれば説明できなくなる(火山移動)のと同様、古小笠原陸塊も今の小笠原と相同のもの(位置や性格)である必要はない(小笠原は“大洋島”ではないという解釈もある)。小笠原固有種が、ずっと小笠原にいたとは限らない。連綿と連なる、幾つもの時代、幾つもの陸塊を、移り住んできた、という可能性も考えられる
そうであるならば、オガサワラシジミの成立も、ルリシジミそのものから派生したのではなく、ルリシジミがスギタニルリシジミ・ウラジロルリシジミと分化する前の、アリサンルリシジミなどとの共通祖先型に基づくと見做すことも可能である。
♂交尾器から見ても近縁各種とは確実な安定差があり、どこか(日本本土や中国大陸)から島に移って、短い期間(数千年とか数万年とか)の間に特化した集団とはとても考え難い。
一般に種の成立には数百万年(人類もそうだ)を擁し、その間の隔離と変化に伴って、独自の形質・性質が齎される(蝶の場合交尾器の形状に如実に表現される)。
もっとも、それだけが種の成立過程とは限らないという見方もできるかも知れない。日浦勇氏や川副昭人氏らの談話会の際、「種のごとく振る舞うことで成立した種」という話題が出てきたことがあった。数100万年かかるであろう必須手順をすっ飛ばして、ショートカットで実質的な種としての機能を獲得(仮免許のようなもの)、後付けで紛いなき種としての独立性を確立するものもあれば、何らかのきっかけで元の集団に収斂していくものもある。遺伝的に異なるのに、形態・形質差が無いという、いわゆる隠蔽種の存在も、その一端なのかも知れない。
まあ、数100万年でも数万年でも、人間の認識からは想像もつかないような修羅場が繰り返されてきたであろうことは想像に難くはない。それをこの10数年間に為された、外敵の出現や、一度や二度の気候変動で説明できるなどとするのは、大いなる思い上がりだと思う。何か、もっと大きな、人知の想像を遥かに上回る、複雑多様な要因が関与している、と考えた方が妥当ではないだろうか。
ただ、21世紀に切り替わった頃に、様々な生物の種や地域個体群が、突然消滅してしまっている、という事実が確かにある。何かがある。むろんその“何か”に人類の行動が大きく関わっているのだろうことは間違いないだろうが、それだけ、と言うわけでも無さそうに思う。自然の摂理のようなもの。
例えば、余りに非科学的な話になるが、進化とか環境とかが関与しない、種としての“賞味期限”のようなもの。人類を含めた地球上の様々な現存生物の多くが、種として数100万年の歴史があるとすれば、(いつか来る大地震みたいに)一斉に滅亡期に突入する、ということがあっても良いのかも知れない。
それに関して気になるのは(もちろん非科学的です)ルリシジミが最近目立って減ってはいまいか?だとすれば、オガサワラシジミを含めたルリシジミ一族(注:分類学上のtribeではない)の生命力が衰えつつある、と言うことである(そんなことはまずないと思うけれど)。
オガサワラシジミの絶滅に至った理由を、研究者や専門家たちが、理路整然と説明している。外敵の増加、環境や気候の急変。それを人為調節できなかったことを悔やんでいる。さらに人為による繁殖の失敗、それは慎重になり過ぎたからで、もっと早くに現地放出し再移入を測るべきだった、等々。
いろんな意味で、違うんではないかい? と思う。
ここで、いつも例に出すジョニー・ティロットソン絡みの話を。
1964年春、ビートルズがアメリカに上陸来襲(ブリティシュインベーション)して、それまで主役を占めていたポップアイドル的歌手たちは見事に一斉駆逐されてしまった。一面ではその通りだと思う。でも、よくよく検討分析すれば、タイムラグがあるのですね。
正確には、ビートルズによって駆逐されたのではなく、ビートルズたち新勢力のブレイクを齎した時期(および要因)と、旧アイドルたちの衰退を齎した時期(および時期)が、重なっている、ということ。旧いのに飽きてきて、新しいのに飛びついた。
(エルヴィス登場後の1957年からビートルズ上陸前の1962年頃にブレイクした)ビートルズ上陸直近のアイドル的歌手たちのヒット曲の推移を仔細にチェックしたところ、彼らが第一線で継続して活動していたのは、5~6年から長くて7年間。57‐58年スタート組は、概ねビートルズ来襲1~2年前の62~63年には勢力が衰え、ビートルズ旋風の時点では既に第一線から退いていた。一方、61-62年スタート組は、ビートルズ来襲の1~2年後の65~66年頃まで第一線で活躍。ということは、もともと全盛期は限られていたわけで、ブリティッシュインベーションに関わらず賞味期限切れで退場したのに過ぎなかった。
もちろん、新世代台頭と、旧世代衰退は、同じ要因に発しているわけで、大いに関係はあるのだけれど、
直接の関わりはない。
帰化種の繁栄と、在来種衰退も、同じ構図。要因は(非常に複雑多様だが)共通し、むろん一部直接の影響(捕食など)もあるだろうけれど、問題はそれだけではない。
ちなみに、アイドル的歌手達も、表舞台からは退いたけれど、その後もそれぞれの自分たちの音楽を地道に発表し続けているわけで、ある意味むしろ健全な状況に戻ったとみることもできる。
オガサワラシジミも、島のどこかに生き残っていると思う。そしてそれ(大発生などせずに細々と暮らしていること)が健全な状態なのかも知れない。
数100万年、あらゆる修羅場を乗り越えて生き続けてきたネイティブ固有種は、少々の気候変化とか人為攪乱とかで滅びてしまうなど、ヤワではない。
巷に言われるグリーンアノール(侵入者代表)とオガサワラシジミ(ネイティブ代表)の関係にしても、両者に早い時期から接してきた僕にすれば、違和感満載である。間違っている、とは言わない。でもそれだけが正解ではない、と。
オガサワラシジミの幼虫は、見事に食草の蕾に溶け込んでいる。確実にそこにいると確信していなければ見つけ出すことは出来ない(それでもアノールの眼からは逃れられないのかも知れぬが)。あるいは、幼虫は信じがたいほどの相当なスピードで移動する。常に草上にいるとは限らず、むしろ地上に潜んでいる時間の方が長いくらいである。
あと、僕の感触では(保護団体などが躍起になって植生回復に取り組んでいる)オオバシマムラサキは、
本来のメイン(ベーシックな)食草ではないと思う。
ルリシジミの食草が様々な科の植物に跨るように、オガサワラシジミの食草も多岐に亘っているのではないだろうか? 蕾、花芽、若葉など肉厚部分を食することのできる、クス科を含む様々な植物。とりあえず、本命とされているムラサキシキブ属に絞っても、半ば雑草的性格を持つ陽樹のオオバシマムラサキではなく、急斜面の、かつより閉ざされた林間ギャップなどに生じるシマムラサキのほうが、本来のオガサワラシジミ生育地と重なっているのではないだろうか?
探しに行きたいですね。(国家権力による実質的妨害を含む)ハードルも予想されるし、先に記したように、理由はともかく、オガサワラシジミと時を同じくして21世紀に切り替わった前後に、各地で多くの生物が絶滅してしまっている、という現実に対する一抹の不安はあるにしても。
幾らかの予算。カメラ。体力。それらさえ整えば、一度時間をかけて島中のチェックを行いたい。もし見つけたら、公式な発表はしない(少なくとも正確な場所は教えない)。国や都や権威機関が、保護とか種保全とかの名目で、飼育個体群の放蝶などに取り組み始めたら、目も当てられないし、、、。
・・・・・・・・・・・・
現時点で、オガサワラシジミの(70年代を中心とした)撮影写真は、ほとんど見つけ出せていません。手許にあるのは、「小笠原緑の島の進化論」に使用した写真(のコピー)と、90年代の写真の一部、それにクオリティの著しく低い、ストロボ使用(失敗例)の数枚の写真、それに加えて1-6回の冒頭で紹介した写真を再掲。

オガワワラシジミ(再掲)
母島猪熊谷Sep.23,1993(以下4枚同じ)

オガサワラシジミ

オガサワラシジミ(再掲)

オガサワラシジミ

オガサワラシジミ
オガサワラシジミ

母島Jul.18,1978

オガサワラシジミ(再掲)
母島Aug.1,1988

オガサワラシジミ(再掲)
母島Aug.1,1988

オガサワラシジミ
撮影データ確認中

オガサワラシジミ
撮影データ確認中

オガサワラシジミ
最初の渡島時(11976年)に撮影した写真の一枚(撮影日データ確認中)

オガサワラシジミ
極めてクオリティの低い♂開翅写真
母島乳房山南面尾根上に左右の谷から吹き上がってくる
Jul.6,1976

オガサワラシジミ(再掲)
母島乳房山Jul.6,1977

オガサワラシジミ
父島中央山Aug.8,1979

オガサワラシジミ(再掲)
父島中央山Aug.8,1979