








哺乳類科学という雑誌の哺乳類進化研究の特集から、2番目の化石研究についての総説ー西岡佑一郎,楠橋直,高井正成.哺乳類科学,60(2):251-267,2020「哺乳類の化石記録と白亜紀/ 古第三紀境界前後における初期進化」ーを読んでみました。
生物の分岐分類学や古生物学における重要な用語が二つあります。一つがクラウン・クレードで、ある分類群を構成する子孫とそれらの祖先のうち派生形質を共有する種を含めた単系統群(クレード)を意味します。私はこれを、原生する多くの種を派生したおおもととなった群と解釈しました(下図でいうと、真獣類、後獣類、アウストラロスフェニダ類)。もう一つがステム・グループで、クラウン・クレード以外の祖先、つまり同分類群に含まれる、あるいは近縁であると考えられるが、明確な共有派生形質をもたない種を意味します。私はこれを、原生する多くの種を派生せずにどん詰まりで絶滅してしまった群と解釈しました(下図でいうと、真獣類、後獣類、アウストらロスフェニダ類以外の群)。
我々人類を含む有胎盤類のクラウン・クレード、すなわち現生哺乳類の系統(現生目)が,白亜紀(中生代)と古第三紀(新生代)との境界(=K/Pg 境界、約6600 万年前)にある絶滅イベントの前に放散したのか、後に放散したのかという問題は哺乳類学における一大研究トピックであり、今なお論争の真っ只中であるとのことです。恐竜全盛期には哺乳類はこそこそ隠れながら生活していたのが、恐竜が絶滅したことで生活の場が広がり、様々な土地、生活様式に適応して種が多様化したと考えるのが理にかなっていそうです。
近年は、現生種の遺伝子情報と化石を含む形態情報を統合して解析し、複数の化石記録を参照して各系統の分岐年代を推定する「ビックデータ分析」が主流で、O’Leary et al.(2013)によるScience誌(本総説の引用文献では間違ってNature誌と書かれている)に掲載された研究が有名だということで、本総説では彼女らの論文がひんぱんに引用されていることからも、この分野における一つの到達点であり、今後に向けた参照点としての重要性がうかがわれます。彼女らは各目レベルのクラウン・クレードの最古となる化石記録に基づき推定分岐年代を算出した結果、有胎盤類の起源と放散はK/Pg 境界後に起きたと結論づけているそうです。さらに、最近の研究では、有胎盤類のクラウン・クレード(異節類やローラシア獣類など)の起源は白亜紀(中生代)で、目レベルでの放散は暁新世(新生代)に起きた可能性が示されていて、現状ではそれが化石記録と分子系統の折り合いをつける最適な解釈だろうとしています(下図では、真獣類(=有胎盤類)のラインが白亜紀にやや太くなって、新生代でさらに太くなっていることで示されています)。

分岐年代の推定には、分類が明らかでかつ産出年代の定まっている化石種が用いられます。しかし、古生物学的な証拠は断片的で、化石の分類や年代値が再評価されることも頻繁にあるため、「最古の化石記録」がどのような裏付けに基づいており、どの程度信頼できるデータなのかという点を明確にしておくことが現代の哺乳類学研究者にとって重要な情報だとしています。つまり、古生物学的な証拠は信頼性が重要ではあるものの、時代を経ることで再評価される可能性もあるので、その時点における暫定的な結果として捉えておいたほうがいいのかなとも感じました。
ここから先は、各論に入って非常に細かな記述が続くので、私なりにポイントをピックアップしていきます。
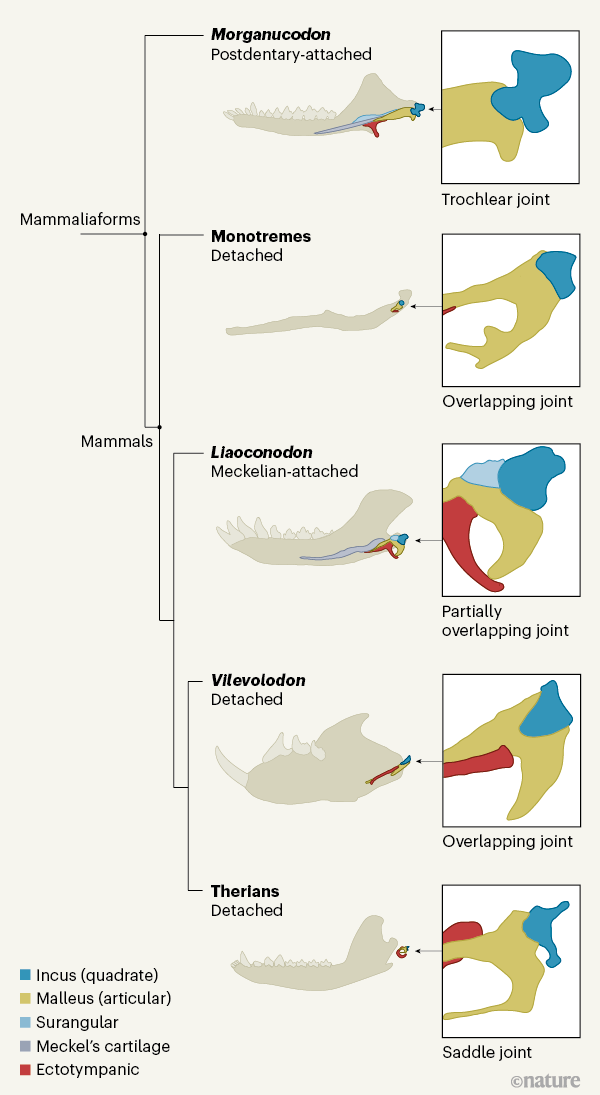
・クラウン・クレードとしての哺乳類とそれに近縁なグループを含めて哺乳形類と称します。現時点での最古の化石記録から、哺乳形類は2億3000万年前までにキノドン類から分岐し、そこから哺乳類が現れたと考えられていますが、中生代の三畳紀からジュラ紀にかけてのどの年代だったかは不明だそうです。ハラミヤ類は哺乳類の多丘歯類の姉妹群で、三畳紀から出ているので、この時代には哺乳類が出現していたという説と、ハラミヤ類は哺乳形類のステム・グループであるという説(上図の位置)があります。このことから、哺乳類の出現した年代がいつなのかは、ハラミヤ類がどこに分類されるかにかかっているところがあります。先日の「哺乳類進化研究アップデート No.4ー哺乳類への中耳の進化」で紹介した論文では、中耳の形態からハラミヤ類は哺乳類に含まれると主張していました。
・中生代の哺乳形類は主に臼歯の形態に基づき、上図のように分類されています。
・単孔類はアウストラロスフェニダ類に含まれます。現在の単孔類はオーストラリアにのみ生息していますが、アウストラロスフェニダ類の中生代の化石はアフリカ、南米、オーストラリアで知られています。
・後獣類(有袋類)と真獣類(有胎盤類)の分岐がいつ起こったかは、哺乳類進化研究の重要なテーマの一つです。近年、その分岐はジュラ紀だという説が出ていますが、まだ証拠不十分で確定していないとのことです。有胎盤類の起源・多様化の時期に関しては3つの仮説、短期結合モデル、長期結合モデル、爆発的放散モデルがあります。分子系統学は、短期結合モデルか長期結合モデルを支持していて、有胎盤類は白亜紀(約1億年前)に出現し、目レベルでの多様化は短期間(短期結合モデル)、または1億年前から6600万年前までの長期間(長期結合モデル)で起きたと考えています。一方、古生物学は、有胎盤類の化石が中生代から発見されていないというO’Leary et al.(2013)などの主張から、爆発的放散モデルを支持していて、K/Pg 境界(6600万年前)の直後に原生目が出現したと考えています。しかしながら、ゲノムの変化の後に形態的特徴が定着するため、分岐年代は分子生物学による推定値よりも化石記録に基づく年代値の方が当然若くなるということも考えられています。
・我々が入っている有胎盤類の真主齧類の中の霊長目について、分子系統学においては白亜紀の約8500 万年前あたりに分岐したとされています。また、霊長目が出現した場所としては、古生物学者は北半球のテチス海周辺(現在のカリブ海~地中海~東南アジアに及ぶ地域)、分子生物学者は見は南半球の大陸(アフリカか南米)と考えています。
以上をまとめると、化石による物的証拠と分子系統樹という計算による予測の間では、それぞれの分類群の分岐の年代において様々な食い違いがあることが分かりました。また気づいた点として、①本総説において真主齧類とローラシア獣類をまとめて北方獣類とする記載は出てこないことから、北方獣類という分類群は分子系統学に特有なもののようであること、②本総説には約100報の論文が引用されていますが、日本人によって報告された論文は1報もなく、この分野の研究ー古生物学的手法による哺乳類進化の研究ーは日本においては低調な状況であることを感じました。


 </picture>
</picture>