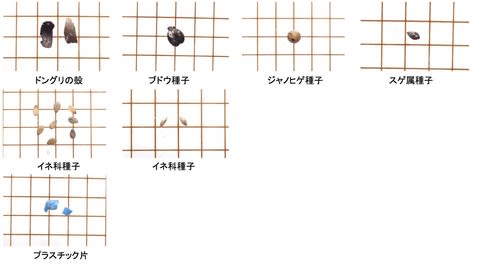
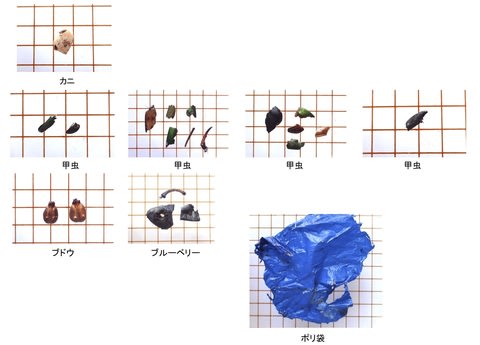
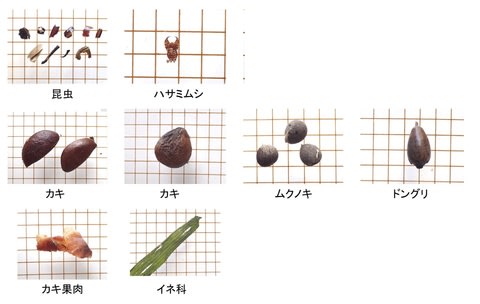
シカ生息地で代表的な群落を選び、新鮮なシカの糞を10の糞塊からそれぞれ10粒拾った。糞サンプルは0.5mm間隔のフルイ上で水洗し、残った植物片を顕微鏡でポイント枠法で分析した。カウント数は200以上とした。
結果
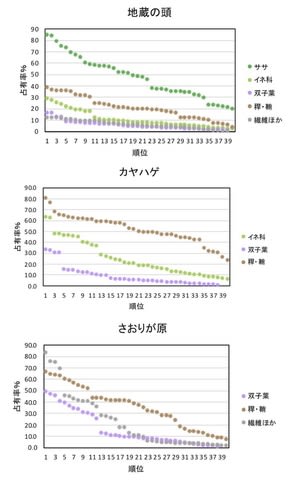
食物は次の9つのカテゴリーに分けた。ササ、イネ科、双子葉(双子葉植物の葉で、顕微鏡下では網目の葉脈やモザイク状の表皮細胞が認められる)、枯葉(不透明な葉脈だけが残り、表皮細胞はない)、果実・種子、繊維(木質繊維と樹皮)、稈(イネ科の茎)、その他(透過性はあるが識別不能、不透過なため識別不能など)である。
各季節の垂直変異
中ライン:冬には垂直的な変異は非常に小さかった。どこでもササと双子葉(多くは常緑広葉樹の葉)、繊維が多かった。
春になるとササは減少し、稈が非常に多くなった。高地ではイネ科が多く、繊維も多かったが、中標高と低地では稈が非常に多く、繊維は少なく、ササが10%前後あるという違いがあった。
夏には低地でサンプルが得られず、垂直比較ができるのは中ラインの高地と中標高のみであった。春と同様、高地でイネ科・稈が多く、中標高では繊維が多かった。ササは1年で最も少なくなった。
秋には垂直的な違いがあり、高地では春、夏と同様イネ科・稈が多かった。中標高ではササが増え、低地では春よりも稈が減り、繊維が増えた。双子葉は冬よりは少なかった。


図2a. 中ラインでの各季節の垂直変異
東ライン:冬は垂直の違いがあり、高地ではササが多く、中標高ではササがさらに多く、低地ではササが少なく、双子葉、繊維が多かった。春も同様の垂直変異があり、高地でイネ科・稈が増え、中標高ではササが大きい値を維持し、繊維と稈が増えた。低地ではササと双子葉がへり、稈が増えた。夏には高地しかサンプルがなく、イネ科が非常に多かった。秋も垂直的な違いが大きく、高地はイネ科が多く、中標高はイネ科・稈が多く、低地は双子葉と繊維が多かった。

図2b. 東ラインでの各季節の垂直変異
以上、場所による違いがあったが、概して高地ではイネ科・稈が多く、低地では双子葉が多い傾向があり、中ラインでは変異が小さかった。ただし東ラインでは垂直的な変異が大きい傾向があった。
場所ごとの季節変化
中ライン:低地では冬にササと双子葉(常緑広葉樹の葉が多い)が多く、春には稈が非常に多くなり、秋は春と似ていたが、繊維が多くなった。冬よりは稈が多かった。中標高では冬と秋がササと双子葉が多いという点で共通していた。春にはここでも稈が非常に多くなり、夏には繊維が非常に多くなった。高地では冬だけがササと双子葉が多く、それ以外はイネ科・稈が多く、春と夏は繊維が多かった。

図3a. 中ラインでのシカの糞組成の季節変化
東ライン
東Lでは冬と秋しかサンプルがない。これらの組成は似ており、双子葉が20%程度あり繊維が30%程度あった。東Mでは冬と春はササが非常に多く40-60%を占めた。夏はサンプルがなく、秋はササは少なく、イネ科が30%台あった。東Hでは冬にササが非常に多かったが、そのほかの季節では少なく、代わりにイネ科が多かった。ただし春には繊維が40%近くを占めた。
このように東ラインでは高さによる違いも、季節による違いも大きい傾向があった。

図3b.東ラインでのシカの糞組成の季節変化
西:冬と秋に双子葉と繊維が多く、春と夏にはイネ科・稈が多くなった。春には繊維も多かった。

図3c.西でのシカの糞組成の季節変化季節変化
以上、季節変化を見ると、冬の糞にはササがある場所ではササと常緑広葉樹の葉が多く、春になるとイネ科にシフトし、イネ科の葉と稈が多くなった。
考察(未完)
丹沢は山体が大きく、地形的にも植生的にも多様であるから、シカの食性も単純には論じられない。今回の分析でも統一的な傾向を読み取るのは困難であった。それでも、これまでにない精度のサンプリングを行ったことにより、従来知られていなかったことがいくつか指摘できた。そのことを中ラインを中心に考える。
<季節変化>
冬には、どの標高でもササと双子葉植物(常緑広葉樹)が多かった。これらの植物の冬の栄養価はよくわからないが、占有率だけを見ると、冬の緑葉が他の季節よりも多いという意外な結果であった。
春にはイネ科が多くなった。シカの採食影響を受けた場合、双子葉草本と比較するとイネ科は分げつにより再生できるから、生き残りやすい。実際、丹沢では低木や、シカが好まない種を除けば、双子葉草本が減少して、相対的にイネ科が生き残ったとされる(田村, 2013)。ただしイネ科の葉は少なく、稈が非常に多かった。これは生育初期のイネ科の葉は消化率が良いために食べたもののうち稈が多く残るからと考えられる。ただし、高地の糞では葉と稈が同じ程度であったが、繊維が多いという違いがあった。
注目されるのは、夏に糞が得にくかったことである。特に低地では全く発見できなかった。この理由はよくわからないが、食物が豊富になるとシカが低地から山に登って行くのかもしれない。ただ、中標高でも高地でも、夏には糞は発見しにくく、発見された糞にしばしばエンマコガネ系の糞虫が見られたから、糞虫によって分解されるために、糞が少なかったことは間違いない。
夏に糞が得られた中標高では繊維が60%以上を占め、高地でも稈と繊維で70%を占めており、葉は少なかった。また糞から量は多くないが枯葉も検出された。このことは、丹沢のシカは夏でも非常に劣悪な食糧事情にあることを強く示唆する。雨が降った場合、生きた植物はいうまでもなく、枯葉があるだけでも雨滴が地面を直接打たないため、土壌流失を抑制する効果があるが、これらが少なくなると土壌流失が大きくなることが知られている(石川ほか, 2007, 畢力格図ほか, 2013)。したがって、夏にシカが枝や枯葉を食べることが、土壌流失に影響している可能性が大きい。丹沢のシカが夏でも枝や枯葉を食べていることの発見は本分析の大きな成果であった。
さらに意外なことに、シカが夏よりも秋の方が良い食物を食べていることであり、低地と中標高では糞中にササと双子葉がやや多かった。高地では葉の占有率は夏と変わらず、繊維が減り、稈が非常に多くなり、ササはなかった。この理由はよくわからないが、中標高でササが増えたのは草本類が枯れ始めてササの必要性が大きくなったからかもしれない。
<塔ノ岳での季節変化>
1994年に塔ノ岳で人馴れしたシカの採食行動を観察した調査例がある(三谷ほか, 2005)。それによると、そのシカは冬にササ、春にイネ科のヒメノガリヤス、夏に緑化工で吹き付けた牧草、秋に枯葉をよく食べたという。本分析の結果はこれとよく対応している。すなわち塔ノ岳(東H)では冬にササ、春にイネ科・稈・繊維、夏にイネ科・稈、秋にイネ科・枯葉が多かった(図3b)。観察では稈や繊維は評価されないし、1994年当時とは植生も同じではないことを考えれば、非常によい対応を示しているといえよう。
<垂直分布>
糞組成の垂直分布をみると、高地でイネ科が多かった。一般に尾根は乾燥しがちであり、また風を受けやすいので、尾根の樹木は斜面や沢に比較して風倒被害を受けやすい。通常であれば、風倒木によってできた森林ギャップは後継樹によって補完されてゆくが、丹沢の場合、シカの強い採食によってそれができず、草原が広がっている。鈴木・山根(2015)は空中写真を解析して、1970年代に比較して2000年代は丹沢の尾根でブナ林が減少して草原が54%も拡大していることを明らかにした。尾根の草原には双子葉草本類もあるが、この草原もシカの採食影響を受けて、再生力のあるイネ科が相対的に多く、そのことが高地でシカの糞にイネ科が多いことの理由になっていると思われる。
<他地域との比較>
夏にシカの糞中に葉が少なかったことについて、他の場所のシカと比較してみたい。岩手県五葉山のシカの場合、代表例として山地帯のミズナラ林(標高1150m)を取り上げると、一年を通じてミヤコザサが重要で夏から秋まで70%台を占め、春には実に89.0%に達した。夏には他の単子葉が21.6%を占め、葉が95%を占めた(Takatsuki, 1986)。
山梨県の乙女高原のシカの場合もミヤコザサが多いが、大きな季節変化を示し、多いのは11月から4月までで、ほかの植物が枯れるため常緑のササの利用度が高くなることがわかった(Takahashi et al, 2013)。初夏にはササ利用は少なくなり、稈が50%程度を占め、秋にはイネ科が20%、稈が30%程度であった。
これらの事例から葉とその他に分けて、葉が占める割合だけを取り出してみた(図4)。すると五葉山のシカの糞は一年中ほとんど葉だけに占められていることがよくわかる。これに対して乙女高原では6月には葉が30%を下回り、7月には40%ほどになるなど、乙女高原でも初夏には葉が少なくなることがわかる。この時期に多くなるのは丹沢と同じく稈であった。丹沢ではそれ以上に葉が少ないが、秋(10月)には乙女高原とほぼ同レベルであった。

図4a.岩手県五葉山、山梨県乙女高原、丹沢(中ライン、中標高)のシカの糞に占める葉の占有率の季節変化。五葉山(Takatsuki, 1986)と乙女高原(Takahashi et al., 2013)は元のデータから計算し直して作図した。
したがって、ミヤコザサの豊富な五葉山(表日光でも同様である)とは大きく違うが、ササがさほど多くない乙女高原とはある程度、似たパターンをとった。ただ、それに比べても丹沢では春と夏は大幅に葉が少ないことが確認できた。
次に食物状態の劣悪さの指標として、糞中の繊維の占有率を比較した。これによると、五葉山でも乙女高原でも繊維は一年を通じて5%未満であったが、丹沢では秋と冬には20%台となり、特に注目されるのは夏に60%以上になったことである。ただし春には4.5%に止まった。このことから、丹沢のシカにおける葉の占有率は乙女高原とやや似ていたが、繊維占有率では大きく違うことが確認された。

図4b.岩手県五葉山、山梨県乙女高原、丹沢(中ライン、中標高)のシカの糞に占める繊維の占有率の季節変化。五葉山(Takatsuki, 1986)と乙女高原(Takahashi et al., 2013)は元のデータから計算し直して作図した。
丹沢では長年のシカの影響でササは少なくなっているが、それでも冬の食物としては重要度が高い。そのことはササへの影響が非常に強く出ることを予測させる。今後、さらにササが減れば、冬季のシカの栄養状態に悪影響を与えるようになるであろう。また、夏にも十分に葉を利用できない状況にあるようだった。
この分析により、丹沢においては、シカの糞に、春から秋にかけての植物生育期においてさえ、葉が少量しか検出されないことがわかった。このことは、現状の丹沢のシカが劣悪な食物環境に生きることを強いられていることを示唆する。したがって、これまで以上に、植生のモニタリング、シカの頭数管理、特にその質的特性(妊娠率や蓄積脂肪など)を把握しながら適切なシカ管理を進めることが重要であろう。そのためにも、糞分析による食性解明は有益な情報を提供するであろう。
文献
石川芳治・白木克繁・戸田浩人・片岡史子・鈴木雅一・内山佳美 . 2007. 丹沢堂平地区のシカによる林床植生衰退地における降雨と土壌侵食量. 関東森林研究 58: 131‒132.
国立公園協会.1964. 丹沢大山学術調査報告書,神奈川県,横浜
鈴木 透・山根 正伸. 2013.空中写真からわかるブナ林の衰退. 森林科学, 67: 6-9.
高槻成紀. 2006.「シカの生態誌」、東京大学出版会
田村 淳. 2015.神奈川県丹沢山地におけるシカ問題の歴史と森林保全対策.水利科学, 333: 52-66.
畢力格図, 石川 芳治, 白木 克繁, 若原 妙子, 海 虎, 内山 佳美. 2013.丹沢堂平地区のシカによる林床植生衰退地における降雨量, 降雨係数および地表流流出量と土壌侵食量との関係. 日本森林学会誌, 95: 163-172.
古林賢恒・丸山直樹. 1977. 丹沢山塊札掛におけるシカの食性. 哺乳動物学雑誌, 57: 5-62.
古林賢恒・山根正伸. 1997. 丹沢山地長尾根での森林皆伐後のニホンジカとスズタケの変動.野生生物保護 2 :195-204.
三谷奈保・山根正伸・羽山伸一・古林賢恒. 2005. ニホンジカ (Cervu snippon)の採食行動からみた緑化工の保全生態学的影響 -- 神奈川県丹沢山地塔ノ岳での一事例. 保全生態学研究, 10: 53-61.
山根正伸. 2012.シカの管理─奥山に登ったシカ.「丹沢の自然再生」、木平勇吉・勝山輝男・田村 淳・山根正伸・羽山伸一・糸長浩司・原慶太郎・谷川 潔編,日本林 業調査会,東京),283-295.
Takahashi, K., A. Uehara and S. Takatsuki. 2013. Food habits of sika deer at Otome Highland, Yamanashi, with reference to Sasa nipponica. Mammal Study, 38: 231-234.
Takatsuki, S. 1986. Food habits of Sika deer on Mt. Goyo. Ecological Research, 1: 119-128.
Yamada, H. and S. Takatsuki, 2014. Effects of deer grazing on vegetation and ground-dwelling insects in a larch forest in Okutama, western Tokyo. International Journal of Forestry Research, 2015, Article ID 687506, 9 pp.