都市における鳥類による種子散布の一断面
■はじめに
森林の動態はきわめて複雑であり,その解明は容易ではない(例えば中静 1994).樹冠を形成する樹木は果実を生産して更新を図る.風散布をする植物もあれば,動物による散布をする植物もある.堅果類は動物によって貯食され,多肉果は動物に食べられることで散布される.複数の植物種がそれぞれの果実を生産し,複数種の動物によって移動されるため,その動きは複雑になる.多肉果の場合,しばしば鳥類が重要な種子散布者であり,ある多肉果樹木が結実すると,その果実が落下すると同時に,飛来した鳥類が採餌のために滞在して消化した種子を排泄したり,口から吐き出したりする.その種子には当該樹の種子だけでなく,その樹木に飛来する前に採餌した樹木の果実も含まれる.多肉果の散布では鳥類が有力な散布者ではあるが(小南 1993:Kominami et al. 2003),テン(ホンドテン)やタヌキなども散布するから(高槻 2017, 2018),種子の動きはさらに複雑である.このように森林の多肉果の散布動態は極めて複雑であると推察される.
現象そのものが複雑である上に,実際に森林で調査をする上でも樹冠が隣接したり,重なり合っていれば,どの樹冠から落下したかの区別が困難であるし,低木・草本が生育し,リターなどもあるため,種子の発見・回収にも困難が伴う.
このような複雑な現象を捉えるために大規模で長期的な調査を行うことで重要な成果も得られているが(例えばNaoe et al. 2018),同時に小規模な事例の蓄積も重要であろう.この点,都市緑地は単純な系であり,孤立木を対象にできるし,しばしば樹下がコンクリート舗装面であり,種子を発見・採集しやすい.そのような例として,唐沢(1978)は1970年代に都市鳥類を調べる過程で鳥類による種子散布を詳細に調べたが,その後は調査事例が少ない(ただし故選・森本 2002など).
以上の背景から本調査の目的は複雑な森林生態系における種子散布の理解の参考にするために,都市緑地の単純な系を利用することで,鳥類による種子散布の実態を解明することを目的とした.
■方法
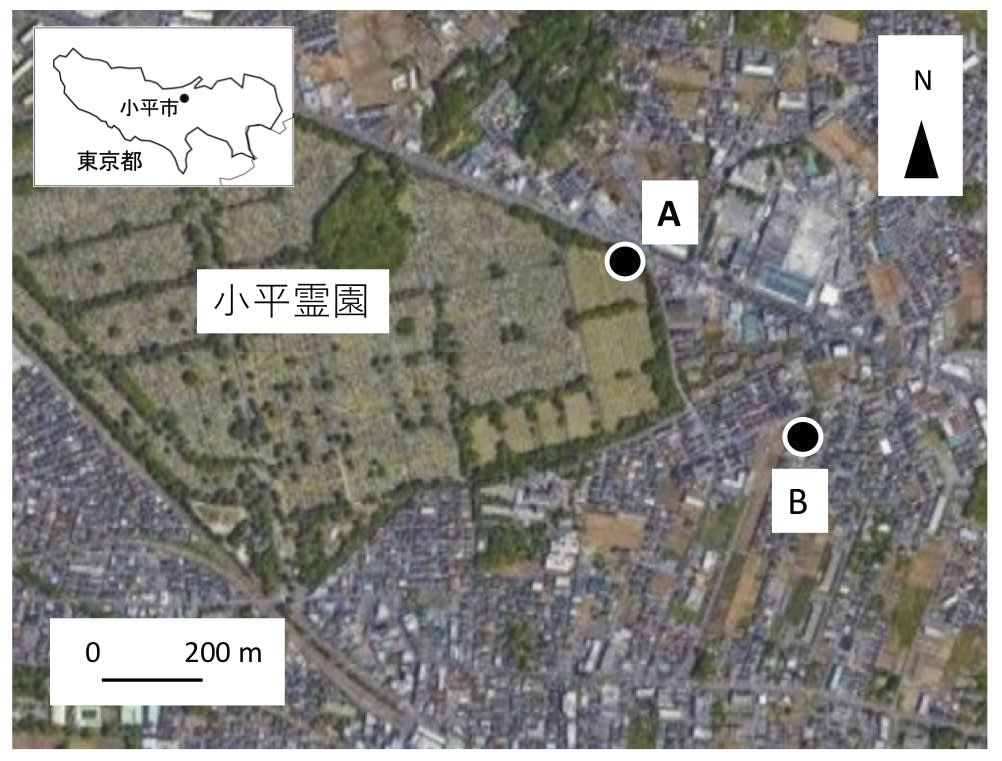
調査は東京都の多摩地区北部にある小平市で行なった(図1).小平市は人口約20万人で,農地,公園などの緑地が比較的多い.
図1. 調査地の地図.ただし調査地Cは範囲外.Google Earthをもとに作図.
調査地A:小平霊園のトウネズミモチの木の下(北緯35°44’ ,東経139°29’)で種子を採集した.小平霊園は65ha,1948年に開園され,面積は65haであり,園内にはケヤキ,ソメイヨシノ,トウネズミモチ,アカマツなどが植栽されている.調べたトウネズミモチの下にコンクリートの歩道がある.

小平霊園のトウネズミモチ(2018年1月)
調査地B:小平市大沼公民館のクロガネモチの下(北緯35°44’ ,139°29’)で種子を採集した.樹下はアスファルト舗装されている.
小平市大沼公民館のクロガネモチ(2018年1月)
調査地C:青梅街道駅近くのある駐車場の電線の下(北緯35°43’ ,139°28’)で種子を採集した.駐車場はアスファルト舗装されている.
種子は調査地Aと調査地Bは2018年1月4日,調査地Cは1月7日に,幅1m,長さ5mの帯状区を調査地A, Bでは樹冠下に,調査地Cでは「止まり木」の下にとって丁寧に拾いあげた後で,箒で掃きとった.被食されないで落下した果実も多数あったが,これらは対象外とした.
■結果と考察
落下種子集団
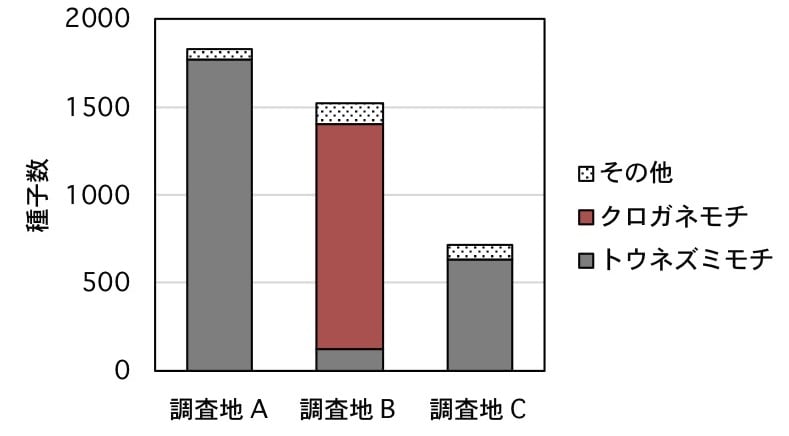
被食種子は母樹由来の種子が多く,調査地Aでは合計1845粒で,トウネズミモチが大半の1770粒(95.9%)を占めた.調査地Bでは合計1535粒で,クロガネモチの種子が1277(83.2%)を占めた(図2).「止まり木」の下である調査地Cの種子数は727粒と少なく,トウネズミモチが638粒(87.8%)を占めた.
回収された種子
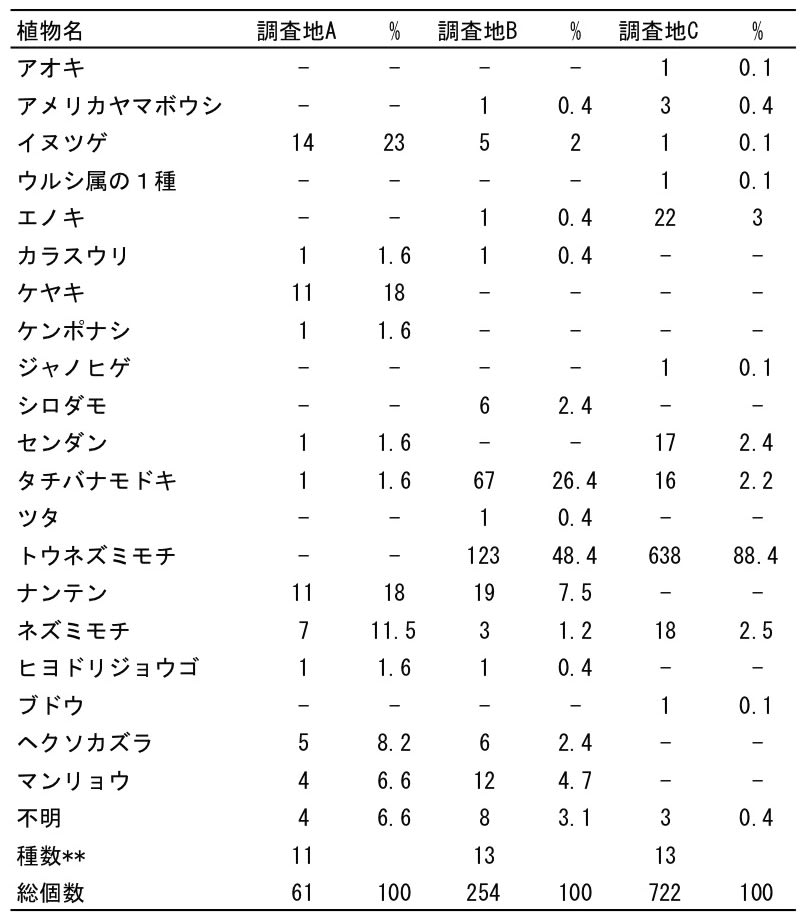
回数された種子は20種が同定され,3種は識別不能種だった(表1).数が多かったのはトウネズミモチとクロガネモチで,この2種は前述のように母樹からの落下が大半であった.
図2. 小平市の3カ所において鳥類に散布された種子の母樹由来とその他の数.母樹は調査地Aがトウネズミモチ,調査地Bがクロガネモチ,調査地Cは母樹なし.
外部から持ち込まれた種子としては,調査地Aではイヌツゲ,ケヤキ,ナンテン,調査地Bではトウネズミモチ,タチバナモドキ,調査地Cではトウネズミモチ,エノキなどが多かった(表1).
表1 外部から持ち込まれた種子の数
果実のタイプ
20種の果実のタイプをみると,ほとんどは「多肉果」であり,堅果はケヤキ,乾果はウルシ属(Toxicodendron)の1種にすぎなかった.ケヤキは果実をつけた枝が数枚の葉を利用して風散布するから(星野 1990),回収されたケヤキ種子は風散布したものかもしれない.鳥類がケヤキを食べるという記録は著者が検索した限り発見できなかった.なおニホンザルはケヤキの果実をよく食べる(辻・中川 2017).
ケヤキの枝先.果実は枝先全体で風で散布される
乾果であるウルシ属の1種はヤマハゼであるかハゼノキであるか識別が困難であった(付図1.1-8).ウルシ属の仲間は種子の外面に脂質に富んだ物質があって鳥類が好むことが知られている(ヌルデRhus javanica var. chinensis:桜谷2001,ウルシ科:佐藤・酒井 2001;上田・福居 1992,ヤマウルシ:原田・上田 2005;桜谷 2001).
そのほかの多くは多肉果であり,鳥類が食べやすい直径10mm以内の球形のものが多かった(唐沢 1978;濱尾ほか2010).ただし,カラスウリは果実が例外的に大きく,鳥類は果肉をついばむことは知られていたが,著者が検索した限り,鳥類が種子を食べたという情報は一般書(多田 2017)以外,発見できなかった.なお,哺乳類では,タヌキでは報告例がないが,テンでは九州で同属のキカラスウリとモミジカラスウリが食べられたことが報告されている(荒井ほか 2003;足立ほか 2016).本調査によって鳥類が種子を散布することが確認された.
カラスウリ果実
ケンポナシは形態学的には果肉でなく果柄部分が肥厚したものであるが,生態学的には「多肉果」として機能している.哺乳類はケンポナシをよく採食するが(高槻 2018),鳥類による採食の記録はこれが初めてだと思われる.
ケンポナシの「果実」
またジャノヒゲの種皮は青くつやがあって多肉果のように見える.
ジャノヒゲの種子
野生植物・栽培植物
回収された20種を野生植物であるか栽培植物であるかで分けると,栽培種は5種に過ぎなかったが,9種は野生植物だが植栽されることが多く,植栽されない野生植物は6種であった(表1).そして種子数は野生種が調査地Aでは77.0%であり,栽培される野生種が調査地Bで75.2%,調査地Cで93.5%であり,いずれにおいても野生種の種子数は0.1から13.1%に過ぎなかった.このことは調査地が都市環境であることを反映していると考えられる(表2).
表2. 種子の類型と種子数
持ち込まれた種子
調査地Aには11種が外部から持ち込まれた(表1).とくに多い種はなく,イヌツゲ(23.0%),ケヤキ(18.0%),ナンテン(18.0%)が上位を占めた.ケヤキは風散布の可能性が大きい.調査地Bでは13種の持ち込み種子があり,種子数はトウネズミモチ(48.4%)とタチバナモドキ(26.4%)が多かった.調査地Cでは持ち込み種子種数は10とさほど違いはなかったが,種子数の大半はトウネズミモチ(88.4%)であり強い偏りがあった.
まとめ
都市緑地の単純な系を利用することで都市の多肉果をつける樹木や「止まり木」の下には,10種あまりの種子が鳥によって持ち込まれることがわかった.3カ所に共通していたのはトウネズミモチが多いということであった.東京都で行われた調査(唐沢 1978)でも同様の結果が得られている.トウネズミモチは明治時代初期に導入された中国原産の樹木で,東京圏の公園,庭,墓地,道路などに植栽されている.吉永・亀山(2001)によれば,トウネズミモチは1960年代から東京の公園などに大量に植栽され,1970年代以降,実生による繁殖によって分布を拡大しているという.
本調査では鳥類の調査はしていないが,断片的な観察ではヒヨドリが多かった.唐沢(1978)も東京都内での調査でヒヨドリが最も多かったとしているし,吉永・亀山(2002)もトウネズミモチに飛来して果実を採食したのはヒヨドリが最も多かったとしている.したがって,現在の東京圏のヒヨドリは冬の食物として外来樹であるトウネズミモチに大きく依存し,種子散布をしていることになる.京都府で鳥類の種子散布に注目した樹木調査をした故選・森本(2002)も京都府の鳥類ではヒヨドリが多く,その種子散布によってトウネズミモチは分布を拡大しつつあることを示した.このことは都市緑地の植栽樹によって鳥類の食性と種子散布が大きく影響されることを示唆し,さらに他の都市でもこの種の調査が行われることが期待される.
調査したのは単純な系とはいえ,母樹由来の種子を除いた種子集団は調査地AとBで大きな違いがあり,その説明は容易ではない.散布者である鳥類の種類が違う可能性があるが,この点は不明である.鳥の種によって食べる果実は違い(唐沢 1978;平田ほか2009;小南・青木 2015),今後は鳥類調査を含めた調査が必要であろう.またこの調査は冬だけしか行わなかったが,原田・上田(2005),田中・佐野(2013)がおこなったように,そのほかの季節も調査することが望ましい.
■摘要
1.東京郊外の小平市の3カ所で鳥類による種子散布を調べたところ,トウネズミモチとクロガネモチの樹下にはそれぞれの母樹由来の種子が多かったが,それ以外に持ち込まれた種子が10種あまりあった..
2. 母樹由来以外の外部からの持ち込み種子は,トウネズミモチの樹下ではイヌツゲ(23.0%),ナンテン(18.0%)などが,クロガネモチの樹下ではトウネズミモチ(48.4%)が多かった.
3.「止まり木」になっている電線の下にはトウネズミモチの種子が88.4%と多く,全体としてトウネズミモチが多かった.
4. 鳥類が散布する種子はトウネズミモチを代表として栽培植物が多かった.
5. 都市緑地の単純な系と舗装地面を利用することで鳥類散布の一面が明瞭に示されることを指摘した.
付図1.1 鳥類によって散布された種子.1.イヌツゲ,2.クロガネモチ,3.ヒヨドリジョウゴ,4.トウネズミモチ,5.ネズミモチ,6.ヘクソカズラ,7.アオキ,8.ウルシ属.,9.マンリョウ,10.アメリカヤマボウシ(ハナミズキ),11.ツタ ,12.ブドウ属.格子間隔は5mm.
付図1.2 鳥類によって散布された種子.13.センダン,14.カラスウリ,15.エノキ,16.ケンポナシ,17.ケヤキ,18.タチバナモドキ ,19.ナンテン,20.ジャノヒゲ ,21.シロダモ.格子間隔は5mm.