Long-term changes in food habits of deer and habitat vegetation: 25-year monitoring on a small island
Seiki Takatsuki
Ecological Research, 2021: 1-10.
https://doi.org/10.2108/zs210055 こちら
1975年から2000年までの25年間、シカが高密度で生息する金華山島のススキ群落とシバ群落で植生とシカの糞組成をモニタリングした。大型草食獣による植生変化がたの大型草食獣に影響与える研究はあるが、自らの食性に与える影響は知られていない。また長期的な植生変化の調査はあるが、草食獣の食性の長期調査はない。調査開始から最初の10年間にススキ群落はシバ群落に入れ替わり、これに伴ってシカの食性もススキ、アズマネザサ、シバからほぼシバだけに変化した。シバ群落は強い採食圧により維持されるが、これにはシバの生産特性と高温多湿な日本の気候によるものと考えた。
草食獣による植生への影響は地形や気候が植生に及ぼす影響に比べれば短期間に変化するものであり、草食獣の密度によって変化する。例えば、乾燥地で過放牧になれば植生荒廃が起きることが知られているし(Fernandez–Gimenes, 2000; Hilker et al., 2014)、島にシカが導入されて植物を食べ尽くすという事例も知られている(Klein 1968; Leader-Williams 1988)。しかし多くの場合はそこまで影響は強くなく、植生は動的に変動する。大型草食獣が植物を食べることは生息地を変形することでもある。その場合、ある動物の採食が植生を変化させ、そのことが別の草食獣に影響を与えることがある。セレンゲティではヌーが高い草を食べることがガゼルにプラスになる(Bell 1970)。一方、ヌーの採食によって草丈が低くなるとトピはそれを利用できなくなるというマイナスの影響もある(Murray and Illius 2000)。これは草食獣が資源あるいはハビタットを変化させることによる他種への間接効果と言える。このような変化は同じ年に起きる季節的な変化であり毎年繰り返されるものである。一方、草食獣による植生の変化は長い時間をかけて起きるものがある。Rooney and Weigman (2004)は五大湖地方の森林を50年前と比較し、オジロジカにより草本類が減少し、シカが食べないシダや再生力のあるイネ科が増えたことを示した。そのようなシカの影響を示した長期の植生変化の調査はあるが(Whitney 1984; Van Deelen et al. 1996; Barrett et al. 2006)、シカの食性を長期的に調べた例は知られていない。
北日本の金華山島はシカが50頭/km2程度の高密度で生息しており、シカ密度は森林で低く、草原で高い(高槻1983)。草原の一つに島の西部の緩斜面があり、1970年代には草丈が1-2mのススキ草原であった。シカの密度が200/km2程度であり、1976年の調査ではシカはススキやアズマネザサ、シバ食べていた(Takatsuki 1980)。この場所は次第にススキが減少し、ススキの株の間に匍匐生の「芝生」であるシバ群落が入り込むようになった。そして1990年台前半になるとススキが非常に少なくシバ群落が広がるようになった。シカはシバをよく食べることが観察されるから、シカの食性が大きく変化した可能性がある。
そこで本調査はシカ生息地においてシカの食物が生息地の食性を採食することの結果としていかに変化するかを25年という長期調査によって示すことを目的とした。
調査地
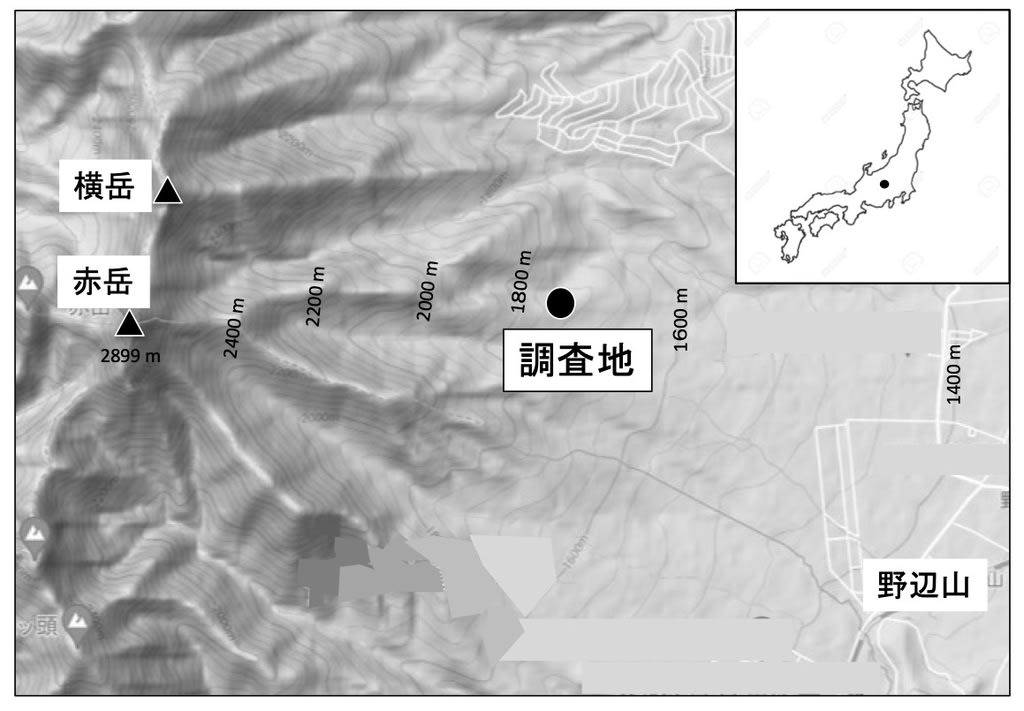
金華山島は本州の太平洋側、牡鹿半島の先端約500m、北緯38度17分、東経141度34分に位置し、面積は992haである(図1)。
図1 調査地の位置図
宗教上の理由で野生動物は保護され、シカは500頭程度が生息している。植生は大半は森林だが、一部に森林がない場所があり、ススキ群落が見られる(吉井・吉岡1949)。島の西部に神社(調査地1)があり、ここでは餌付けをしているためにシカの密度が300/haと高く、採食圧が強いため、シバ群落が広がっている(図2)。この景観は25年間変わらなかった。
図2. 調査地1、2の景観の推移
(調査地2)この神社の北側に緩斜面があり、ススキ群落がある。ここには1980年代に30頭程度のシカ(密度100/ha)がいた。ここは1970年代から1980年台前半には草丈1mくらいのススキ群落で、アザマネザサも多かった(Takatsuki 1977, 図3)。調査地2の一角に面積20m2ほどのシバ群落があった。ここにはメギやアザミもある。これらの量は変わらなかった。
この島には200年前からシカがいたことがわかっている(宮城県史編集委員会, 1960)。頭数は1966年以来直接目撃で追跡されており、当時は40-45頭/km2出会った(Ito 1968)。その後増加して1983年には70頭/km2になり、1984年に厳冬があって大量死が起きた(Takatsuki et al., 1991, 1994)。約半数のシカが餓死した。3年で50-60に回復し、1996年まで安定していたが、1997に再び大量死が起きた。その後再び3,4年で回復した。この変動と自然死はシカ頭数は環境収容力ギリギリであることを示す。調査地1と2の密度は300 頭/km2、100 頭/km2である。この密度は調査期間中変化しなかったが、調査地2では1967には46頭/km2であった(Ito 1968)。Ito(1968)は調査地2のシカが調査地1のシカに攻撃行動をとったことを観察している。しかし本調査が始まった1970年代には調査地1のシカはしばしば調査地2に入った。これは1970年代の経済復興後訪問客が急増し、牡鹿半島に道路がついたことによる。訪問客はシカに餌付けをし、増加して調査地2の密度は倍増した。本調査はこの増加後に始まった。
方法
主要種の被度と草丈
調査地2において1975.8/7, 1984. 9/13, 1988.10/13, 1992年9/18に南北30m間隔の東西のライン10本をとり、10歩間隔で1 m x 1 mのプロットをとり、ススキ、アズマネザサ、シバの被度を記録した。プロット数は約400点であり、被度は10%刻みの目測を記録した。草丈は各種について100個体をランダムに選び、1cmの精度で測定した。
糞分析
調査地1と調査地2において1985年8月17日、1988年8月28日、1991年8月26日にシカのフン20粒を10の糞塊から採集し(Campbell and Johnson 1983; Homolka 1993; Klein and Bay 1994)、顕微鏡によりポイント枠法(Stewart 1967)で分析した。計数したポイント数は200以上とした。食物は、ササ、ススキ、シバ、他のイネ科、他の単子葉、双子葉、カン、繊維、その他に分けた。これらを1975年の分析結果(Takatsuki 1980)と比較した。これら3つの年代を通じて一度でも占有率が10%を超えた植物を「主要食物」とし、Kruskal-Wallis検定(Steel-Dwass事後検定)した。主要食物はススキ、シバ、アズマネザサ、その他のイネ科、カンであった。「不明」も10%を超えたが、これは異質な内容を含むので主要食物にはしなかった。
調査地1と調査地2の各年代の糞組成をもとにWhittakerの百分率組成を算出した。
結果
被度
1975年以降のススキ、アザマネザサ、シバの被度の推移を図5に示した。1975にはプロットの64.6%はススキの被度が20%未満であり、被度が60%以上は7.9%に過ぎなかった。被度はその後減少し、1992年には98.6%が被度20%未満になった。
アズマネザサはススキと似た変化を示した。被度は1975年には48.2%のプロットが20%以上であったが、1992年にはわずか2.4%となった。
シバは上記2種とは対照的に1975年にはプロットの63.6%が被度20%未満であったが、そのような被度の小さいプロットは1992年には19.8%に減少し、被度80%以上のプロットは1975年にはわずか5.6%であったが、1992年には48.1%まで増加した。
図5. 調査地2におけるススキ(A)、アズマネザサ(B)、シバ(C)の被度(%)分布の推移
草丈
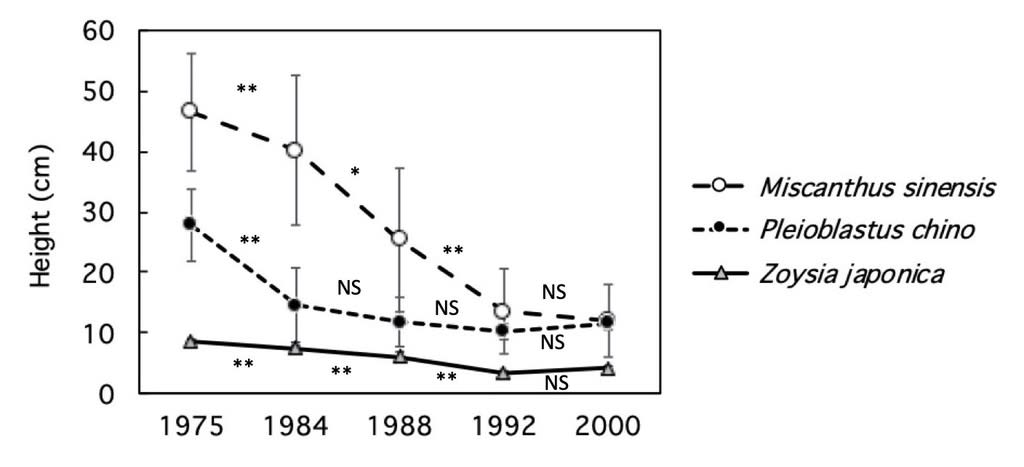
ササ、シバ、ススキの草丈の平均値は3種とも減少したが、特にススキの減少が大きかった(図6)。ススキは1974年から1984年にかけては有意差がなかったが、1984年から1988年と、1984年から1992年には有意に低下した。ササは1975年から1985年の減少が大きく有意に低下し、その後は漸減したが、有意差はなかった。シバはもともと低いので、各年代で有意差があったが(最大値(1975, 8.6cm)と最小値(1992, 3.3cm)の差でも5.2cmに過ぎなかった。
図6. 調査地2におけるススキ、アズマネザサ、シバのと草丈(cm)年推移。バーは標準偏差。
糞組成
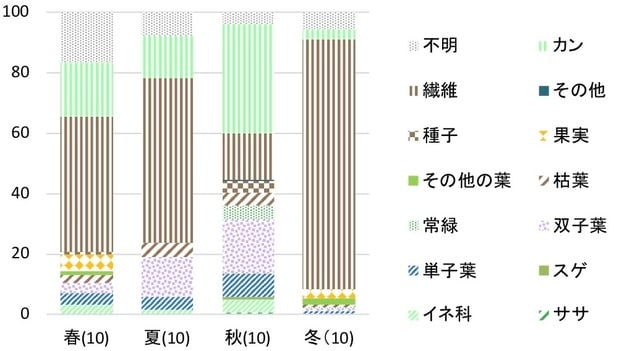
1976, 1985, 1991の夏(8月)のフン組成(付表1)のうち主要種の占有率を図7に示した。調査地1ではシバが60-80%を占め、そのほかの食物カテゴリーとしては、1976年のササと1988年のカンだけが10%以上になった。
ササは1976に11.0%であったが、その後全く出現しなくなった。ススキは常に10%未満であり、1976に5.1%であったが、1985に1.1%有意に減少し、1985から1988は微増し、1988から1991は有意差がなかった。これら2種に対してシバは非常に大きな占有率であった。1976に58.4%であり、1985には75.0%に有意に増加し、1985から1988は有意差はなかったが、1988から1991は有意に減少した。その他のイネ科は常に10%未満であった。1976に8.7%であり、その後有意差がなかった。カンは1976に5.0%であり、1985に微増し、その後は有意差がなかった。このように調査地1では1970年代にシバが58%であり、その後は60%以上で全体にシバが優占していた。
図7. 調査地1, 2におけるシカの糞中の主要食物の占有率の推移
調査地2では1976年から1985年の間に大きな変化があった(図7)。
ササとススキは1976年から1985年にかけて減少した。ササは1976に25.0%を占め、1985には有意に減少した。1985から1988も有意に減少したが、1988から1991は有意差はなかった。ススキは1976に12.0%であったが、1985に4.1%有意に減少し、1985から1988は有意差はなかったが、1988から1991に有意に微増した。これら2種に対してシバは1976年以降増加した。1976に25.8%であったが、1985には59.8%に有意に増加したが、その後は有意差はなかった。その他のイネ科は10%前後を占めた。1976に14.2%であり、その後多少の増減をした。カンは1976に4.9%であり、その後有意差はなかった。このように調査地2では1970年代と1980年代にササとススキが減ってシバが増えるという大きな変化があった。
糞組成の類似度
調査地1と調査地2の糞組成をWhittakerの類似度百分率で比較すると図8のようになった。1976年のPSは66.5%であったが、その後増加して、1988年と1991年には85%前後になった。この内容は調査地2のシカ糞においてシバが増加したことにある。
図8. 各時期の調査地1と調査地2のフン組成のPS
考察
1970年代から2000年にいたる25年間のモニタリングにより、シカ高密度な草原でtussock型からlawn型への植生変化が起き、これに伴いシカの食性もtall grassが減少してシバが優占するようになることが示された。金華山は全体に森林群落であり、シカ密度は森林では低く、草原で高いことが知られている(Takatsuki 1983)。草原の中でも違いがあり、ススキ群落に比べてシバ群落が2倍ほど高密度であった。
ススキ群落とシバ群落は互換性があり、強い採食影響下ではススキ群落がシバ群落になり、逆も真である。これはシバ群落に柵を作った場合、柵内でススキが伸びてシバを庇蔭し、シバが減少することで示される(Ito and Takatsuki, 2005; Takatsuki and Ito 2009)。これはそれぞれの種生態で説明できる。ススキは大型のtussock grassで、中程度の採食で維持されるが(Takatsuki et al., 2009, 2012)、強い採食圧では減少する(Takatsuki et al 2007)。アズマネザサも同様である(Takatsuki, 1980c)。これに対して、シバは匍匐型のlawn grassである、旺盛に茎とtillerをのばす(Ito et al., 2003; Okubo et al, 1977; Otani et al., 2002))。草丈は低いが、生産性は高い(Ito and Takatsuki, 2005)、これによりシバ群落は高密度のシカを支えられる。
本調査でgrazingにより調査地2でススキとササが減ることが示された。ここでのシカ密度は1960年代には50頭 /km2であったが、1970以後に100頭/km2になった(Ito 1968)。これは調査地1でシカが増加して調査地2に入ったからで、その増加は経済復興と牡鹿半島に道路ができたことにより1960年代に観光客が急増したことによる。本調査はその後に始めたことになる。こうしたことから、期間中にススキ群落からシバ 群落への変化は採食によるものであると説明できる。
調査期間中の調査地2での景観変化を見ると(図2)、1970年代はススキが優占し、シバは見られないが、1980年代半ばにはシバが侵入し、1980年代後半も大きな変化はないが、1990年代にはシバ群落の中にススキが少なくなり、メギやアザミが目立つようになった。主要種の被度はススキとササは減少、シバは増加し、その変化は緩慢であった(図3)。また高さも同様であったが、ササの減少は1970年代から1980年代が大きく、その後は安定的であった。シカの糞組成を見ると、1970年代と1980年代で大きな違いがあり、この間にササの減少、シバの増加が見られた(図6)。このことは植生は徐々に変化したが、シカにとっては食物供給に臨界点があって突然大きな変化が起きたと考えられる。つまりススキの被度で20%未満がほぼ半分以上になり、シバの被度20%以上が半分以上になり、ススキの平均草丈が40cm程度になると、シカの食物ではシバが過半量になるということである。シバの被度とススキの被度・高さがさらに減少しても、シカの食物供給としては違いがなかった。この間、シバは草丈を下げたが、シバは現存量が小さくても生産量は大きいから、次々に葉を展開してシカに供給すると考えられる。
このような高密度においても土地の荒廃が起きなかったことは注目される。大型草食獣の採食影響のうち乾燥地の家畜の放牧の場合、家畜の密度が高くなると、荒廃が起き、砂漠化につながることが知られている(Meyer 2006; Liu et al. 2013; Hilker et al. 2014; Gao et al. 2015)。本調査では劣化も侵食も見られなかった。これは日本列島の高温多湿な気候に支えられていると思われる。
Rooney et al. (2004)は五大湖地方で50年前のデータと比較して、オジロジカの採食により草本類が減少し、シカが好まないシダとグラミノイドが増えたことを示した。不嗜好植物はシカにとって利用できないので生息地の劣化になるが、グラミノイドは再生力があるので、持続的に利用されるようになる。金華山でのシバの増加と利用はこれに対比できるかもしれない。シバは盛んに生産し、シカはもっぱらシバを食べた。ところがシバは暗い環境では生産力が下がる(Hirayoshi and Matsumura 1957; Otani et al. 2002)。このためシカによるススキとササの除去はシバにとってプラスになる。つまりシバは採食に耐性があるというより、競争相手を除去してもらっている。
調査期間中調査地1、2では高密度のシカがいたが1984年(Takatsuki et al., 1991; 1994)と1997年(高槻, 2006)に大量死が起きた。捕食者がおらず狩猟も行われていないから環境収容力で制限され、晩冬に自然死が起きる。大量死の後には5年以内で回復した。これはシバ群落の高い生産力に裏付けられている。
そういうシカ-シバ群落関係は各地で知られている(高槻 1980, Takatsuki 1980b, 1982, 1983, 1984, 1985, 1987)。家畜の放牧場でも知られている(Arai and Okubo, 2014; Iwata, 1971)。
有蹄類は多種が群落を変えることで影響を受けることが知られている(Bell, 1970; Murray and Illius, 2000)。この調査は25年間のモニタリングにより、シカが植生を変え、その結果自分の食性を変えた例を示したという点でこれらの研究とは違う。