








さいたま市の浦和商業高校のタヌキの食性
高槻成紀・小林邦夫
摘要
市街地のタヌキの食性分析例として、埼玉県さいたま市の高校の敷地に隣接する雑木の木立ちのタヌキの糞分析を行った。この木立ちは白幡沼という沼に隣接している。サンプルは2021年の1月から12月まで毎月採集し、ポイント枠法で分析した。ここのタヌキの食性は、冬は食物組成が多様で、春はアズマヒキガエルと昆虫が増え、夏はエノキの果実、昆虫、アメリカザリガニが増え、秋はエノキ、ムクノキの果実が優占した。特徴的なこととしてヒキガエルとアメリカザリガニが検出された。このことはタヌキの食性の日和見的性質を示している。検出された種子はカキノキ、ウメ、ビワなどの栽培種を含め7種にすぎず、関東地方の里山環境で検出されるキイチゴ類、クワ属、ヒサカキなどがなく、種数が貧弱であった。さまざまな人工物が検出されたが平均占有率は4.0%にすぎなかった。これらの結果は緑地に乏しい市街地にある学校の敷地とその周辺という、生育する樹木の種数が限定的で、沼に隣接する環境をよく反映していた。
キーワード:アズマヒキガエル、アメリカザリガニ、食性、タヌキ、都市
■ 序
東京周辺のタヌキの食性はかなり明らかになってきた(山本・木下1994酒向ほか2008,手塚・遠藤 2005、Hirasawa et al. 2006、Sakamoto and Takatsuki 2015、Akihito et al. 2016、高槻 2017、Takatsuki et al. 2017、Enomoto et al. 2018、高槻ほか 2018、高槻・釣谷2021)。この地域のタヌキの食性は基本的に果実を主体にしており、特に秋と冬は果実をよく食べる。ただし夏には果実が少なくなるので、食物中に昆虫が多くなり、食物が最も乏しい冬の終わりから早春には鳥や哺乳類の羽毛、毛、骨などが検出されるようになる。これらの調査は主に郊外や山地で行われたが、市街地のものもある。ただし市街地の調査地のうち、皇居(酒向ほか 2008,Akihito et al. 2016)、赤坂御用地(手塚・遠藤 2005)、明治神宮(高槻・釣谷2021)などは都市としては例外的な森林があり、都市的緑地を代表するとはえない。市街地での調査事例としては川崎市(山本・木下1994)と小平市の津田塾大学の事例(高槻 2017)がある。川崎市では果実とともに人工物が非常に多かったが、津田塾大ではそうではなかった。これは家庭ゴミの回収の仕方が変化し、2000年以前にはゴミ回収法が不徹底だったためにタヌキが利用できたが、その後家庭ゴミはボックスなどに入れて回収されるようになったためにタヌキは残飯類などを利用しにくくなったものと考えられる。このように市街地のタヌキの食性分析例は少なく、さらなる分析事例が必要である。
本調査の調査地である浦和商業高校は埼玉県さいたま市にある。ここは交通の要所でもあるために開発が進み、緑地は非常に限定的である。そしてビルや住宅地に囲まれているため、市街地のタヌキの食性調査事例として適している。ただし沼に接している点が特徴的である。
■方法
調査地は旧浦和市、現在のさいたま市南区で(図1)、西側には新幹線、埼京線、東側に東北本線、南側に武蔵野線が走り、線路に囲まれている。
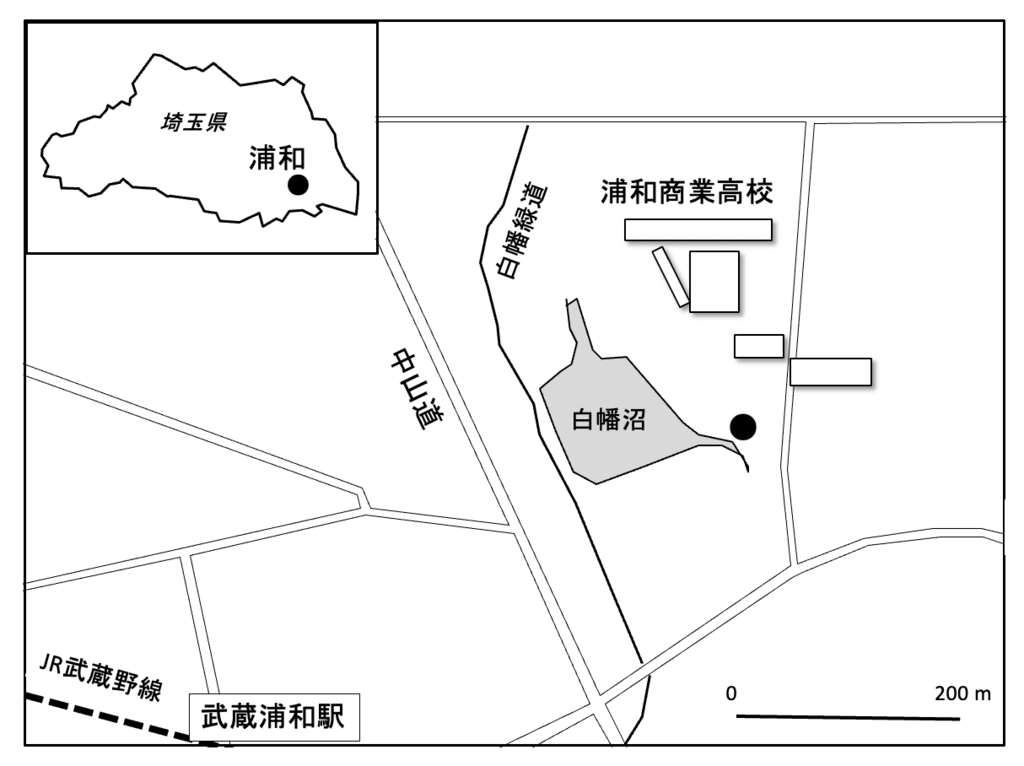
図1. 調査地の地図。●:タヌキの糞採集地
また西側には首都高速大宮線、南側には東京外環自動車道があるなど交通の要所であり、開発が進んでいる。浦和商業高校の西側500 mに武蔵浦和駅があり、その周辺はビル街であるが、浦和商業周辺は学校が多く、住宅地が広がる。農耕地はなく、自然には乏しいが、学校の西側には白幡沼があり、弁天神社の小さな祠があって周囲に木立があり、限定的な緑地となっている(図2)。

図2. 調査地を白幡沼の西側から見た景観
タヌキはこのあたりに生息し、高校生のクラブ活動が終われば明るいうちでも複数の個体が観察される。このように調査地は交通要所にある市街地に囲まれた高校の敷地に隣接する樹林であり、沼に隣接している点が特徴的である。
ため糞はこの樹林内にあり、そこから糞サンプルを回収した。採集にあたっては,糞の大きさ,色,つや,新しさなどから同一個体による1回の排泄と判断されるタヌキの糞数個を1サンプルとし,それを複数採取した.
糞サンプルは0.5 mm間隔のフルイで水洗し,残った内容物を次の15群に類型してポイント枠法(Stewart 1967)で分析した.
哺乳類,鳥類,脊椎動物の骨,昆虫(鞘翅目,直翅目,膜翅目,幼虫など),節足動物(多足類など),甲殻類,その他の動物質,果実,種子,葉(イネ科,スゲ類,単子葉植物,双子葉植物、枯葉),支持組織(繊維、稈など)、植物その他(コケなど)、作物(農作物、栽培果樹)、人工物(輪ゴム,ポリ袋,紙片など),その他.
ポイント枠法では,食物片を1 mm格子つきの枠つきスライドグラス(株式会社ヤガミ,「方眼目盛り付きスライドグラス」)上に広げ,食物片が覆った格子交点のポイント数を百分率表現して占有率とした.1サンプルのポイント数は合計100以上とした.また、食物カテゴリーの占有率を大きい順に並べた線グラフで表現する占有率–順位曲線(高槻ほか 2018)を描いた。
また、採食行動を記録するため調査地内に3台のセンサーカメラ(トレイルカメラPH770ー5S、Abask社)を設置してタヌキの出没や採食行動を記録し、糞分析の参考にした。
■結果
食物内容の季節変化
各食物カテゴリーの占有率(%)の月変化を図3に示した(付表3も参照)。
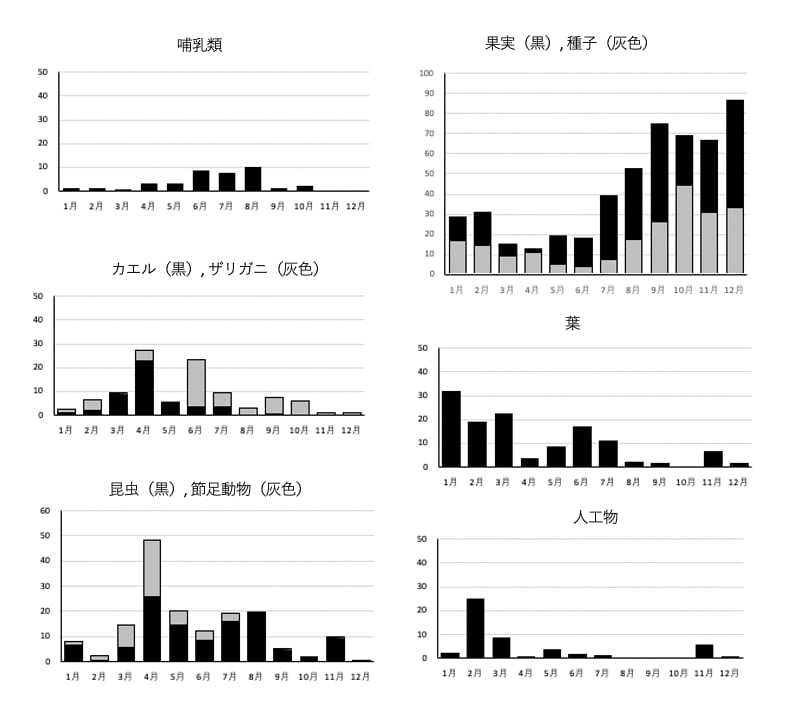
図3. タヌキの糞組成の月変化(2022年)
哺乳類の毛は1月から5月は微量、6-8月には10%前後となり、その後また少なくなった。カエルの骨は3, 4月に多く、特に4月には22%を占め、その後は数%で8月以降はほとんど検出されなかった。ザリガニは5月までは少なかったが6月には19.9%となり、その後10月までは数%から10%以上までの値をとった。
昆虫は4月から8月までほぼ10%以上を占めた。4月の25.8%は最大値だったが、この月には節足動物も最大値(22.7%)をとった。これは昆虫の足や翅と違う細片で昆虫である可能性は大きく、そうであれば合計で50%近くを占め、非常に重要であった。
果実と種子はここのタヌキにとって最重要な食物であった。1,2月はあわせて30%、3-6月には30-40%と少なかったが、7月以降に増加し、9月には最大の75.0%に達し、その後も70%前後を維持し、12月にはさらに増加して80%以上になった。
葉は1-3月に多く20-30%を占めたが、その後は少なかった。
人工物は多い月でも10%未満で、8-10月には全く検出されなかったが、検出されたものは多様で次の通りであった。アルミホイル、プラスチック、ポリ袋、ポリエチレンの袋(いわゆる「レジ袋」を含む)、化学繊維、ゴム製品、輪ゴム、紙、皮革製品、糸、ひも
季節類型
このような結果から、占有率の大きいカテゴリーをもとに季節区分をすると次のようにするのが妥当だと思われた(図4)。
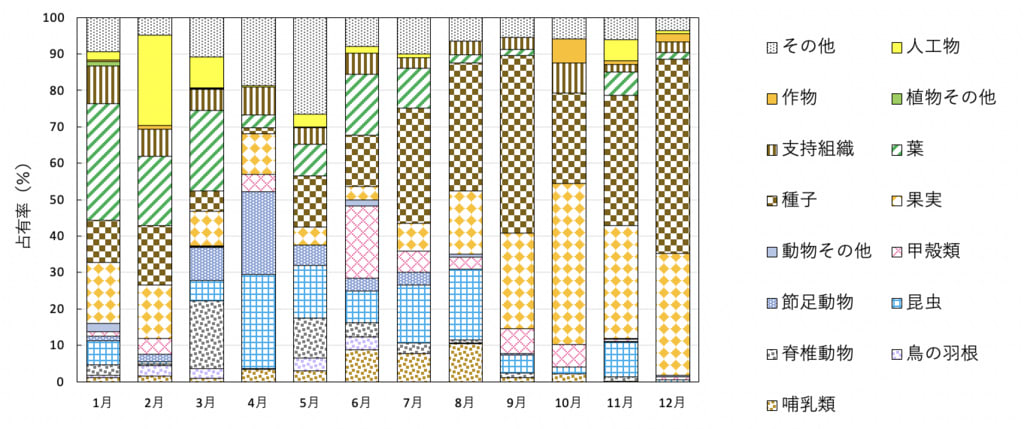
図4. タヌキの糞組成の月変化(2022年)
冬(1-3月):1月、2月は果実・種子がやや多く、葉、農作物、種子などもある程度多く、組成が多様という点で共通し、4月以降とは違った。3月は1月、2月より果実・種子が少ないが、人工物と葉が多い点で2月に似ており、昆虫が少ない点と脊椎動物(アズマヒキガエル)が多い点で4月と違った。3月と隣り合う月との百分率類似度を求めると、2-3月間は51.4%、3-4月間は47.6%で2月の方が大きかったので3月は冬とした。
春(4-6月):昆虫が多く、果実・種子は少ない点で共通していた。
夏(7,8月):昆虫と果実・種子が多い点で共通していた。
秋(9-12月):果実・種子が独占的である点で共通していた。
主要種の占有率-順位曲線
食物カテゴリーごとの占有率-順位曲線を描いた(図5)。

図5. 主要食物カテゴリーの占有率-順位曲線
果実は果肉と果皮(「果実」とする)と種子に加えて、その合計値(「果実合計」とする)を示した。占有率-順位曲線のパターンには安定的に豊富にあって動物がよく利用する高い占有率からなだらかに減少する「高値漸減型」、食物資源が局在するため一部のサンプルが占有率が多く低値も多い「L字型」、低値が少ない「I字型」、供給量は多いが動物が好まないために占有率は小さいが高頻度な「低値高頻度型」などがある(高槻ほか 2018)。「果実合計」は高い値から直線的に減少し、典型的な「高値漸減型」であった。果実、種子はカーブが下にやや窪む形をとった。動物質は最大値が中程度で低頻度なものが多く「I字型」が多かったが、昆虫だけは頻度が高く「L字型」をとった。葉は「L字型」、繊維は低い値である程度高頻度な「低値高頻度型」だった。作物と人工物は最大値が大きいか中程度で低頻度の「I字型」だった。
果実の推移
果皮、果肉からは種の同定は困難なので、種子の占有率を示した(図6)。

図6. 主要種子の占有率の月変化. A:冬から夏に出現した種子、B: 夏以降に出現した種子. 縦軸は一定でない.
センダンが1-4月に出現し、サクラ属が5月、ビワが6月、クワ属が6, 7月に5%前後からそれ未満で検出された(図5A)。エノキとムクノキは多く、ここのタヌキの非常に重要な食物となっていた(図5B)。エノキは1–3月は少なく4月には出現しなくなったが、5月から出現し始め、8月には35%に達し、その後10月には一時的に下がったが、その後再び増加した。ムクノキは1月から5月までは少なく、6–8月には出現しなくなったが、9月以降は20%前後を占め、12月には29.3%に達した。このようにタヌキは季節に応じて推移する果実を利用していた。
■考察
関東平野の大都市の一つである旧浦和市(さいたま市南区)の市街地にある高校一帯に生息するタヌキの食性を糞分析によって調べた。この場所は周辺に農耕地がないこと、昼間は高校生がいるが夕方から夜は無人になること、一般の市街地よりは廃棄物などが得にくいこと、樹木や草本類が限定的であること、白幡沼という沼が隣接し、小規模な樹林があることが特徴的である。
食物はエノキ、ムクノキなどの果実が主要であったが、春にアズマヒキガエルが、夏にアメリカザリガニが食べられる点が特徴的だった。都市、あるいは郊外で果実食傾向があることはこれまでも東京都小平市(高槻2017)、新宿御苑(Enomoto et al. 2018)、東京都日出町(Hirasawa et al. 2006; Sakamoto and Takatsuki 2015)、皇居(酒向ほか 2008; Akihito et al. 2016)、赤坂御用地(手塚・遠藤 2005)、明治神宮(高槻・釣谷2021)などで確認されているが、カエルやザリガニの利用は知られていない。タヌキが日和見的な食性を持つことはこれまでにも指摘さてきたが(山本・木下 1994; Hirasawa et al. 2006; Takatsuki et al. 2021)、この結果もそのことを裏付ける。
エノキとムクノキの果実は特に重要であったが、これらの結実時期は夏以降であるにもかかわらず1–4月の分にも含まれていた。この時期のエノキ、ムクノキ、センダンは前年の夏から秋にかけて結実して落下したものをタヌキが探して食べたものと考えられる。同所的に生息するタヌキとテンの食性を調べた研究では、テンは結実期に果実を食べたが、タヌキは長い期間食べ続けたことが知られている(Takatsuki et al 2017)。カキノキは高校の敷地内にもあり、9月の結実初期には枝についたカキノキの果実を食べようと後肢で立ち上がり、何度か挑戦して最終的に果実に噛みついて枝を折ることに成功して、地面で食べるのがセンサーカメラに撮影された(図7)。

図7. 枝についたカキノキ果実を食べようと後肢立ちになったタヌキ(2022年9月18日).
3月に「脊椎動物」が18.6%を占めたが、その大半はアズマヒキガエルの骨であった。この時期はアズマヒキガエルの繁殖期であり、警戒心がなくなっているためにタヌキが捕食しやすい可能性がある。調査地に設置したセンサーカメラにはヒキガエルを発見し、前肢でコントロールしながら噛みついて引きちぎったシーンが撮影された(図8)。
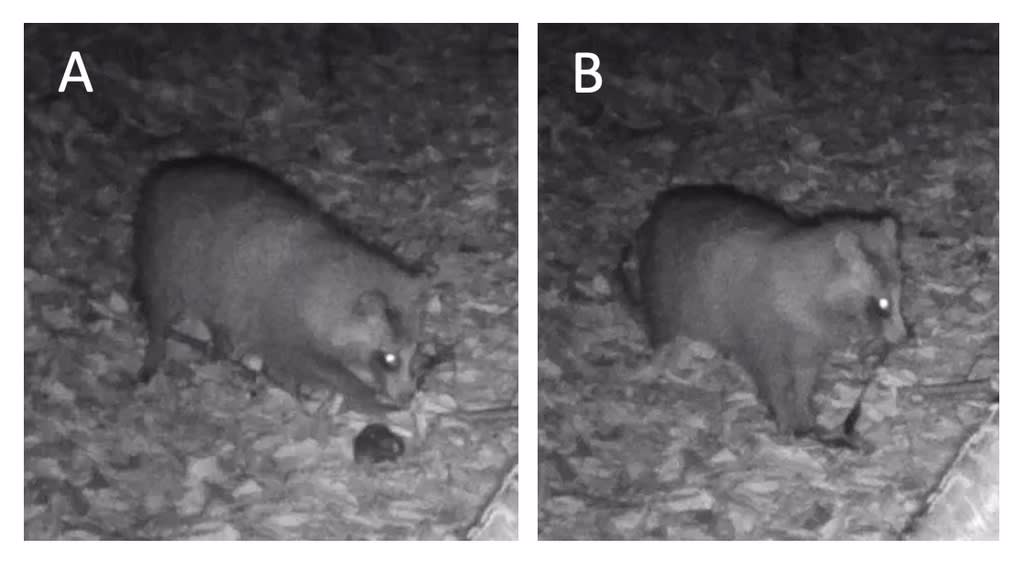
図8. アズマヒキガエルを捕食するタヌキ。A: アズマヒキガエルを見つけて右前肢でコントロールし、B: 引きちぎって食べる(2022年5月12日)
糞中のアメリカザリガニは4月に多く(19.9%)、その後も10月まである程度出現したが、調査地のタヌキによるカエルやザリガニの利用は食物に占める割合は大きくはなかった。特にヒキガエルは利用も短期的であり、アメリカザリガニも出現頻度は35%程度で、高いとはいえない。これらのことから、ここのタヌキは基本的に調査地の木立や高校の敷地内の樹木の果実などを軸に、時々白幡沼を訪問してこれらを利用するという程度であると推察される。しかし、そのことはタヌキが生息地にある食物を順応的に利用してメニューを拡大する潜在力を持っていることを示す好例といえよう。
人工物はアルミホイル、プラスチック片、ポリエチレンの袋、化学繊維、ゴム製品、輪ゴム、包紙、皮革製品、糸、ひもなど多様であったが、出現頻度は全体で23.0%、平均占有率は4.0%にすぎず、食物としての重要度は小さいといえる。人工物が20%以上であったのは果実も昆虫も乏しい2月だけで、これを除けば平均占有率は0.5%にすぎない。これは市街地の緑地に生息するタヌキとしては人工物への依存度が低いといえる。その背景として、高校の敷地であり、タヌキが利用する残飯や捨てられた菓子類などの袋などの供給が一般の公園などより少ないという事情があると思われる。
食性とも関連するが、本調査地のタヌキの状況を考えてみたい。旧浦和市(現在はさいたま市の一部)は、1960年代から1970年代にかけて急激に発展し、人口は敗戦の1945年には94,000人ほどであったが、1960年には倍増して160,000人ほどになり、1988年(平成元年)には410,000人ほどで、敗戦年の4倍以上になった。さいたま市の土地利用の変遷を見ると、1906年には畑と田が広く、両方で60%以上であったが、1969年には田はやや増えたが、畑は8割ほど、樹林は7割ほどに減り、宅地が7割増えた(付表1)。
付表1. 旧浦和市(現さいたま市)の1906年, 1969年, 2006年の土地利用の推移(国土交通省 2012より)

そして2003年になると、田畑を合わせても3割ほどに減少し、樹林は5%になったのに対して宅地は62%に達した。調査地周辺の土地利用の1961年と2022年を取り上げると、1969年には南西部は田畑が広がっていたが、現在は新幹線が通り、武蔵野線もあるので、ビル街となっている(付図1)。
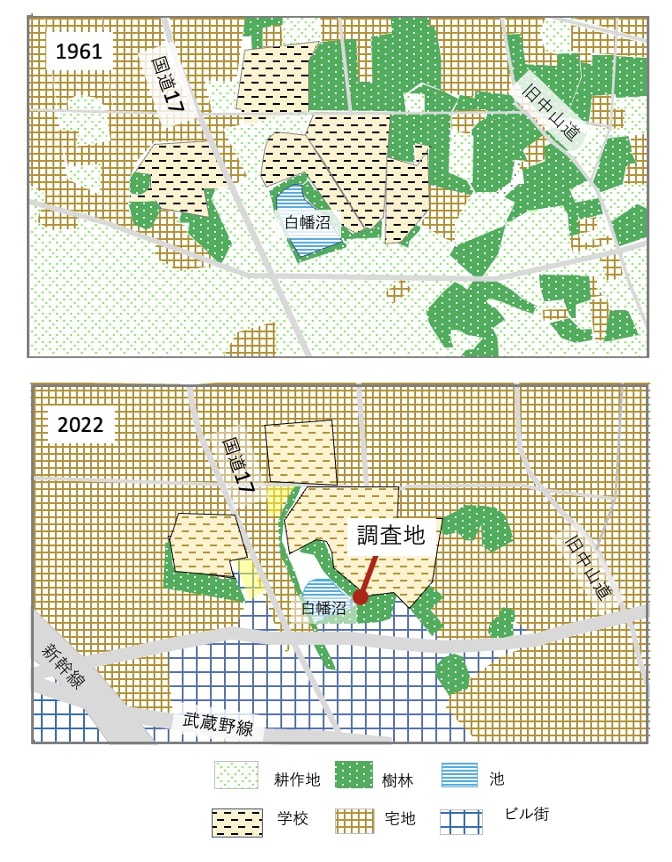
付図1. 調査地周辺の1961年と2022年の土地利用(空中写真より作図)
また北東部に多かった宅地が全体に拡大した。そしてこの範囲では田畑は消滅した。北部から東部には樹林がかなりあったが、この半世紀に減少し、現在はわずかに社寺や公園、学校などにしか残されていない。こうした中で白幡沼と隣接する樹林は貴重な緑地となっている。調査地はこの緑地帯の一角にあり、周辺には樹木のあるような広めの庭のある宅地もある。タヌキはそれらをつなぐように利用している可能性はあるが、人口数万人程度の小都市にあるようなまとまった樹林や農耕地はない。したがっていわば島のように孤立した状態で生息していると考えられる。そのことは食性にも反映しており、タヌキが利用していた食物に農作物と特定できるものは少なく、わずかにコメの籾殻、ソバ、ミカン種子が微量に検出されたにすぎない。調査地周辺に水田やソバ畑、ミカンの果樹園などはなく、何らかの理由で落ちたものを食べたものと思われる。強いて農地的な食物といえばカキノキの果実で、カキノキは高校敷地にもあり、タヌキがそれを食べるところもセンサーカメラに撮影された(図7)。しかしカキノキ果実の利用期間は短く、占有率も小さかった。したがってここのタヌキは農作物をほぼ利用していないといってよい。
そうした中にあって高校の敷地内や周辺にエノキとムクノキが比較的多くあり、タヌキはこれらの果実に依存的である時期が長かった。ただし、タヌキが利用した果実の種数は少なく、これまでのタヌキの食性分析では種子は20種前後検出されることが多かったが、本調査地では7種にすぎず、非常に乏しいといえる。関東地方の里山のタヌキの食物からは、キイチゴ、クワ属、ミズキ、サルナシ、ヒサカキなどが高頻度で検出されるから(Hirasawa et al. 2006; Sakamoto and Takatsuki 2015; 高槻ほか 2020)、これらがなかったことは孤立した市街地にある高校とその周辺という、植物相の単純な環境を反映したものと考えられる。そのことを含め、本事例はタヌキの食性が日和見的であるという見解(Hirasawa et al 2006, Takatsuki et al. 2021)を支持するものであった。
■文献
Akihito, Sako, T., Teduka, M. and Kawada, S. 2016. Long-term trends in food habits of the raccoon dog, Nyctereutes viverrinus, in the imperial palace, Tokyo. Bulletin of National Museum, Natural Science, Series A (Zoology) 42: 143–161.
Enomoto, T., Saito, M. U., Yoshikawa, M. and Kaneko, Y. 2018. Winter diet of the raccoon dog (Nyctereutes procyonoides) in urban parks, central Tokyo. Mammal Study 43: 275–280.
Hirasawa, M., Kanda, E. and Takatsuki, S. 2006. Seasonal food habits of the raccoon dog at a western suburb of Tokyo. Mammal Study 31: 9–14.
Sakamoto, Y. and Takatsuki, S. 2015. Seeds recovered from the droppings at latrines of the raccoon dog (Nyctereutes procyonoides viverrinus): the possibility of seed dispersal. Zoological Science 32: 157–162.
酒向貴子・川田伸一郎・手塚牧人・上杉哲郎・明仁. 2008. 皇居におけるタヌキの食性とその季節変動. 国立科学博物 館研究報告 34: 63–75.
Stewart, D. R. M. 1967. Analysis of plant epidermis in faeces: a technique for studying the food preferences of grazing herbivores. Journal of Applied Ecology 4: 83–111.
Takatsuki, S., Miyaoka, R. and Sugaya, K. 2017. A comparison of food habits between the Japanese marten and the raccoon dog in western Tokyo with reference to fruit use. Zoological Science 35: 68–74.
高槻成紀・岩田 翠・平泉秀樹・平吹喜彦. 2018. 仙台の海岸に生息するタヌキの食性 - 東北地方太平洋沖地震後に復帰し復興事業で生息地が改変された事例 -. 保全生態学研究 23: 155-165.
高槻成紀・山崎 勇・白井聰一. 2020. 東京西部の裏高尾のタヌキの食性―人為的影響の少ない場所での事例―. 哺乳類 科学 60: 85–93.
高槻成紀・高橋和弘・髙田隼人・遠藤嘉甫・安本 唯・野々村 遥・菅谷圭太・宮岡利佐子・箕輪篤志. 2018. 動物の食物組成を読み取るための占有率 − 順位曲線の提案−集団の平均化による情報の消失を避ける工夫 −. 哺乳類科学, 58: 49-62.
Takatsuki, S., M. Inaba, K. Hashigoe, and H. Matsui. 2021. Opportunistic food habits of the raccoon dog – a case study on Suwazaki Peninsula, Shikoku, western Japan. Mammal Study, 46: 25-32.
高槻成紀. 2017. 東京西部にある津田塾大学小平キャンパスにすむタヌキの食性. 人と自然 28: 1–10.
高槻成紀 ・釣谷洋輔. 2021. 明治神宮の杜のタヌキの食性. 鎮 座 百 年 記 念 第 二 次 明 治 神 宮 境 内 総 合 調 査 報 告 書 第 2 報 : 91-100.
手塚牧人・遠藤秀紀. 2005. 赤坂御用地に生息するタヌキの タメフン場利用と食性について. 国立科学博物館専報 39: 35–46.
Whittaker, R. H. 1952. A study of summer foliage insect communities in the Great Smoky Mountains. Ecological Monographs, 22: 1-44.
山本祐治・木下あけみ. 1994. 川崎市におけるホンドタヌキ Nyctereutes procyonoides viverrinus個体群の死亡状況と生命表. 川崎市青少年科学館紀要 5: 35-40.
















※コメント投稿者のブログIDはブログ作成者のみに通知されます