19.9.19
人工林が広がる場所でのシカの食性 – 鳥取県東部の事例 --
高槻成紀(麻布大学いのちの博物館)・永松 大(鳥取大学)
摘要:日本各地でシカが増加し、森林植生に強い影響を与えるとともに林業被害も増加している。スギ人工林が卓越する鳥取県東部の若桜町では過去20年前からシカが急増し、林床植生が貧弱化した。林業被害対策にはメカニズム解析が不可欠で、シカの食性は一つのポイントとなるが、人工林の卓越する場所でのシカの食性は知られていない。植生は、スギ人工林では柵外はコバノイシカグマ以外は非常に乏しかったが、柵内にはチヂミザサ、ススキ、スゲ類などがあった。落葉広葉樹林でも貧弱で、ムラサキシキブなどが散見される程度であったが、柵内ではタケニグサ、ベニバナボロギク、ジュウモンジシダ、ガクウツギ、ニシノホンモンジスゲ、ススキなどがやや多かった。シカの糞分析の結果、シカの糞組成は植物の生育期でも緑葉が20-30%程度しか含まれておらず、繊維や枯葉の占有率が大きいことがわかった。夏に葉の占有率がこれほど小さいのは神奈川県丹沢のシカで知られているだけである。
はじめに
シカ類の増加と森林生態系の問題は世界的に深刻である(McShea and Rappole, 1992, Waller and Alberson, 1997, Côté et al., 2004, Tanentzap et al., 2010)。日本でも過去20年ほどでシカが増加し(++)、森林生態系に利用を及ぼすとともに(Takatsuki, 2009)、林業の被害を起こしている(大井 1999、Akashi N, Nakashizuka T (1999), Ueda et al., 2002, 2003, 尾崎 2004、Honda et al., 2008)。シカ(ニホンジカ)は落葉広葉樹林に生息することが多いが、場所によってはスギやヒノキの人工林が卓越した場所がある。人工林は密植されるため林内が暗く、林床植物は乏しいことが多い。そのため同じ生息密度であっても広葉樹林よりはシカの食物は乏しく、そのことが林業被害とも関連している。林業被害は被害発生のメカニズムを解明することが重要であり(Ueda et a., 2002, 2003, Honda et al., 2008)、シカの密度や食性を解明は重要な情報となる。
シカの食性は北海道から屋久島まで広範に分析され、大体の傾向は把握されているが(Takatsuki, 2009)、まだまだ残された地域も多い。中国地方はその一つで、2000年に山口県のシカで断片的な情報が報告されたにすぎない(Jayasekara and Takatsuki, 2000)。この分析がなされた1990年代後半には中国地方でのシカの生息は限定的であったが、その後、徐々に拡大した。鳥取県においても兵庫県から連続的な分布域が県東部から徐々に拡大傾向にある(鳥取県, 2017)。
このため農林業への被害が大きくなり、鳥取県ではその抑制のために捕獲が進められ、2010年からは3000頭台、2013年以降は4000頭を超えるレベルになっている(鳥取県 2017)。
調査地である若桜町を含む鳥取県東南部で群落調査とシカの糞密度の関係を調査したところ、若桜町はその中でもシカ密度が高く、植物への影響も強いことが示された(川嶋・永松, 2016)。場所によってはもともとはササがあったが、シカによって食べ尽くされ、低木層も貧弱化し、不嗜好植物(シカが嫌って食べない植物)が増えている場所もあった。
シカが高密度になり、植生に強い影響を与えて食物が乏しくなった状況で死体の胃内容物を調べた例では、枯葉が大きな割合を占めていた(Takahashi and Kaji, 2001)。このようにシカは食物供給の状態に応じて食性を変えることができる。人工林が卓越する場所では食物供給状態は良くないと想定され、シカの食性も落葉樹林とは違う可能性がある。しかし、これまで人工林が卓越した場所でのシカの食性は知られていない。
本調査はこのような背景から鳥取県東部の若桜町のシカの現時点での食性を明らかにすることを目的とした。
方法
1)調査地と林床植生
調査地は鳥取県東部の若桜町に選んだ。この地方は伝統的に林業が盛んであり、若桜町の林野率は95%,人工林率は58%に達していて(鳥取県東部農林事務所 2017),「若桜の杉の美林」として知られる。鳥取県東部のスギは常緑であり葉の垂直的厚さがあるために林床は暗く、林床植生は貧弱である。シカの糞は若桜町役場の南側にそびえる鶴尾山(452 m)の山頂周辺,国指定史跡「若桜鬼ヶ城」の隣接地で採集した(図1)。
図1. シカ糞採取地の位置
糞採取した場所はアカマツとコナラの林で、下生えは強いシカの影響を受けて貧弱になっており、場所によっては2m程度のシカの口が届く高さ以下では植物が非常に乏しくなって「ディア・ライン」が認められた(図2)。

図2. シカ糞採集地の景観。下生えは非常に貧弱で、「ディア・ライン」がみられる場所もある。
シカによって影響を受けている林床群落を記述するために、スギ人工林と落葉広葉樹林の林床に1m四方の方形区をランダムに10個とり、方形区内の植物種の被度(%)と高さを記録した。また史跡保護のために設置されている防鹿柵を利用して,比較のために柵設置後10年の柵内でも同様の記録をとった。これらをもとに、バイオマス指数(被度%×高さcm, Takatsuki and Sato 2013)を求めた。そしてバイオマス指数を木本、双子葉草本、グラミノイド(イネ科とカヤツリグサ科の総称)、シダ、つるに分け、草本はさらに大型草本(50cm以上になるもの)と小型草本に分けて比較した。
2)糞分析
シカの糞の採取に際しては1回分の排泄と判断される糞塊から10粒を採集して1サンプルとし、10サンプルを集めた。糞は2018年の5月から2019年1月まで6回採集した。5月は春、6月、7月、9月が夏、11月が秋、1月が冬に対応する。これを光学顕微鏡でポイント枠法(Stewart, 1967++)で分析し、占有率を求めた。ポイント数は200以上とした。
糞中の成分は次の14群とした。
ササの葉、イネ科の葉、スゲの葉、単子葉植物の葉、双子葉植物の葉、常緑広葉樹の葉、枯葉、その他の葉(コケ、シダなど)、果実、種子、その他、木質繊維、稈、不明
枯葉は黒褐色の不透明な葉脈となった落葉樹の葉であり、緑葉は葉肉部もあり、葉脈は半透明であるから区別ができた。中間的なものもあったが、違いが不明瞭なものは緑葉にした。占有率が1回でも10%以上になった食物群を「主要食物」とし、月間の占有率を多重比較した。
結果
1) 林床植生
調査地のスギ人工林と落葉広葉樹林のようすを図3に示した。柵外はどちらの林でも植物が非常に乏しかった。柵内では回復が見られ、スギ人工林ではススキが、落葉広葉樹林では草本類やスゲが目立った。

図3. 林床の景観。A. スギ人工林、B. スギ人工林柵内、C. 落葉広葉樹林、D. 落葉広葉樹林の柵内
スギ人工林と落葉広葉樹林で植被率(%)を見ると、シカの影響を受けている柵外ではスギ林が3.8%、落葉樹林が3.0%といずれも小さく、両者に有意差はなかった(多重比較、クラスカル・ウォリス検定、z = 0.495, P = 0.96)。柵の内外ではいずれも有意差があった(スギ林、z = 3.631, P = 0.002、広葉樹林、z = 3.9621, P < 0.001)。
スギ人工林と落葉広葉樹林で、バイオマス指数を生育型ごとに合計したところ、スギ人工林でも落葉広葉樹林でも柵外では植物が非常に乏しいこと、柵内では大幅に増加することがわかった(図4)。スギ人工林柵外でのバイオマス指数は45.6で柵内の11.7%に過ぎなかった。内訳ではコバノイシカグマが68.3%を占め、目立って多かった。柵内ではスギ人工林ではイネ科が目立って多く、シダ、大型草本がこれに次いだ。
落葉広葉樹林でのバイオマス指数は13.0と少なく、柵内のわずか2.5%に過ぎなかった。内訳ではムラサキシキブなどの木本類が半量程度であった。柵内では大型草本、シダ、木本が多かった。
柵内をスギ林と落葉広葉樹林で比較すると、スギ林でグラミノイド(チヂミザサ、ススキ、ニシノホンモンジスゲ)が多いこと、広葉樹林で大型草本(タケニグサ、ベニバナボログクなど)、木本(ガクウツギ、クロモジなど)、シダ(ジュウモンジシダ)が多い傾向があった。つる植物は柵外にはほとんどなかったが、柵内ではバイオマス指数で10から20程度あった。

図4. スギ林(Con)と落葉広葉樹林(Bro)の柵外と柵内のバイオマス指数
2)シカの糞の組成
主要食物(一度でも10%以上になった食物)にはイネ科の葉、双子葉植物の葉、枯葉、稈、木質繊維の5つであった。その季節変化は以下の通りであり(図5)、主要種を含む各食物群の組成は付表1に示した。
イネ科は季節変化が不明瞭で10%前後を推移したが、9月の3.8%は7月の10.3%より有意に少なかった(多重比較、z=3.403, p=0.009)。
双子葉植物は7月から11月に多い山型を示した(図3)。6月(1.7%)から7月(15.6%)には有意に増加(z=-3.781, p=0.002)、7月(15.6%)から9月(10.7%)に有意に減少(z=3.326, p=0.011)、9月(10.7%)から11月(19.5%)に有意に増加(z=-3.780, p=0.002)、11月(19.5%)から1月(4.6%)に有意に減少(z=3.78, p=0.002)と、増減を繰り返した。
枯葉は春(5月, 17.8%)と夏(9月、25.1%)、秋(11月、19.8%)に20%前後と多かった。しかし隣あう採集月で有意差はなく、有意差があったのは6月(10.0%)と9月(25.1%)の間だけであった(z=-2.948, p=0.038)。
稈は全体に大きな値をとり、夏に多くなる山型をとった(図3)。7月には45.4%と非常に多く、9月(23.4%)に有意に少なくなり(z=3.780, p=0.002)、1月(10.8%)は11月(27.2%)より有意に少なかった(z=3.024, p=0.030)。
木質繊維も全体に多く、特に5月(55.5%)と1月(48.4%)に多くてU字型を示した。6月(36.3%)から7月(4.2%)へは有意に減少(z=3.781, p=0.002)したが、9月(30.9%)には有意に増加し(z=-3.781, p=0.002)、11月(36.3%)には減少(z=3.780, p=0.002)、1月(36.3%)には増加(z=-3.780, p=0.002)と変化した。

図5. 若桜町(鳥取県東部)のシカの主要食物の月変化
考察
群落調査の結果、柵外ではスギ林でも落葉広葉樹林でも植被率が3%程度しかなく、シカの食物という点で言えば、ほとんど食物がないといえる状況であった。特に面積的に広いスギ林ではその乏しい植物のバイオマス指数の半量以上がコガノイシカグマで占められていた。コバノイシカグマはシカが食べず、食べ残された状況にあった。関東地方ではオオバノイノモトソウが同位的な位置にある(高槻、未発表)。
柵内はスギ林でも広葉樹林でも林床のバイオマス指数が大幅に増加し、スギ林では8.6倍に、広葉樹林では37.9倍になった。このこともシカの採食圧の強さを示している。柵内で回復した植物の構成には違いがあったが、これは元々の種組成の違いと、柵設置後の光条件が落葉広葉樹の方が良いことなどが関係していると考えられる。
シカの糞組成において、夏を中心に葉が多く、冬を中心に繊維が多いというパターンは各地のシカの食性で見られるものであった。しかし葉が夏でも30-40%しかなく、9月には16.8%に過ぎなかった点(後述)と、繊維が冬や春には50%前後にもなり、9月でも30.9%になった点は注目に値する。さらに、夏でも枯葉が食べられて、7月には12.9%、9月には25.1%にも達した。これらのことは、本調査地のシカの食物供給状態が劣悪であることを示唆する。
このことを確認するために、岩手県の五葉山(Takatsuki, 1986)、山梨県の乙女高原(Takahashi et al., 2013)、宮城県の金華山(Takatsuki, 1980)、神奈川県の丹沢山地(高槻・梶谷、印刷中)を比較する。五葉山と乙女高原はシカの密度はあまり高くなくて植物が豊富な場所である。金華山はシカの密度が50頭/km2にも達し、植物は強い影響を受けてシカの食物供給は劣悪である。丹沢山地は場所によって密度は違うが場所によっては金華山並みの高密度の場所もあり、シカの影響を長く受けているので植生は貧弱である(村上ほか, 2007)。
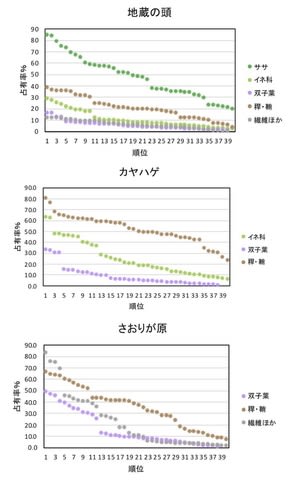
葉の占有率は五葉山が一年を通じて非常に多いが、これはミヤコザサが主体であった(図6)。乙女高原では夏・秋が40%程度、春・冬が50-60%で冬の方がササを食べてやや多くなった。金華山では夏・秋に50-60%と多く、春・冬は20-40%であった。丹沢山地では春と夏が20%程度と少なく、秋・冬にササを食べて40-50%になった。これらに比べると。若桜では春は丹沢と金華山よりは多く、乙女高原程度であり、少ないとは言えなかった。しかし夏は丹沢とともに他の3カ所よりもはるかに少なかった(13.0%)。秋には増加したが5カ所中最少であった(36.3%)。冬も5カ所中最少であった(19.3%)。

図6. 若桜以外4カ所のシカ糞における葉(A)と繊維(B)の占有率。ただし金華山では繊維のデータがない。
繊維は金華山では糞組成を類型するとき「その他」にまとめたので図示できなかった。それ以外の4カ所の比較では、シカの密度が低い五葉山と乙女高原では一年中10%以下と少なかったが、丹沢山地では春以外は多く、若桜では春と冬は4カ所中で最多、夏と秋は丹沢山地に次いだ。
川嶋・永松(2016)は若桜を含むこの地域の植生を調べて、シカによってササが減った場所があると指摘した。シカの糞におけるササの占有率は最も多かった冬でも2.1%に過ぎなかった。しかも、サンプル間のばらつきが大きく、変動係数は繊維が22%であったのに対して、ササは119%であった。このことは調査地ではササが点在していたことと符合する。これは五葉山や乙女高原でササ(ミヤコザサ)が林床全面を覆うのと違い、供給が確実、安定的でないことを意味する。ササは常緑であり、鹿の冬の重要な食物である。このことは五葉山(Takatsuki, 1986)、日光(Takatsuki, 1983)、大台ケ原(Yokoyama et al., 1996)などで示されている。ササの喪失は調査地のシカにとっても林業被害にとっても深刻であろう。
冬は草本類が枯れ、落葉樹は葉を落とすからシカの食物が乏しくなる。そのような状況では植林木への被害が強くなる(Ueda et al. 2002)。この意味で、ササの減少は林業被害につながる可能性がある。
林床群落は非常に貧弱であり、シカの採食影響が強いことを示していた。このことは当然、シカの食性に影響を与えているはずであり、夏でも葉が少なく、枯葉が多いこと、繊維含有率が高いことなどはこのことを反映していると考えられる。
シカの個体数管理は密度を調べて過剰と判断されれば捕獲頭数を決めて駆除がおこなわれる。しかし、シカにとっての資源量である植物量が違えば、密度のもつ意味が違い、植物量の少ない人工林では同じ頭数のシカがいても食物量は少なく、被害が発生する可能性が大きい。その意味でシカ頭数管理は単純に密度ではなく、シカの栄養状態、繁殖率、食性など、シカの状態の質的判断に基づくべきである。この調査の食性分析は、この地方のシカの食料状況が劣悪であることを示した。糞分析方は非侵襲的で比較的簡単であるから応用されることが期待される。
文献
Akashi N, Nakashizuka T (1999) Effects of bark-stripping by sika deer (Cervus nippon) on population dynamics of a mixed forest in Japan. For Ecol Manage 113:75–82
Jayasekara, P. and S. Takatsuki. 2000. Seasonal food habits of a sika deer population in the warm temperate forest of the westernmost part of Honshu, Japan. Ecological Research, 15: 153-157.
川嶋淳史・永松 大. 2016. 鳥取県東部におけるシカの採食による植生の被害状況. 山陰自然史研究, 12: 9-17.
McShea, W. J. and J. H. Rappole. 1997. Herbivores and the ecology of forest understory birds. In The Science of Overabundance: Deer Ecology and Population Management, ed. W. J. McShea, H. B. Underwood and J. H. Rappole. Washington: Smithsonian Institution Press, pp. 268-309.
Stewart, D. R. M. 1967. Analysis of plant epidermis in faeces: a technique for studying the food preferences of grazing herbivores. Journal of Applied Ecology 4: 83–111.
Takahashi,H. and K. Kaji. 2001. Fallen leaves and unpalatable plants as alternative foods for sika deer under food limitation. Ecological Research, 16: 257-262.
Takahashi K, Uehara A, and Takatsuki S. 2013. Food habits of sika deer at Otome Highland, Yamanashi, with reference to Sasa nipponica. Mammal Study, 38: 231-234
Takatsuki, S. 1980. Food habits of Sika deer on Kinkazan Island. Science Report of Tohoku University, Series IV (Biology), 38(1): 7-31.
Takatsuki S. 1986. Food habits of Sika deer on Mt. Goyo. Ecological Research, 1:119-128
Takatsuki, S. 2009. Geographical variations in food habits of sika deer: the northern grazer vs. the southern browser. Sika Deer: Biology and Management of Native and Introduced Populations, (eds. D. R. McCullough, S. Takatsuki and K. Kaji): 231-237. Springer, Tokyo. Elsevier
Takatsuki, S. and T. Kajitani. in press, Food habits of sika deer of Mt. Tanzawa: a case study at a habitat affected by long-term heavy deer grazing. ++ in press
Takatsuki, S. and M. Sato. 2013. Biomass index for the steppe plants of northern Mongolia. Mammal Study, 38: 131-133.
鳥取県. 2017. 鳥取県特定鳥獣(ニホンジカ)管理計画.
鳥取県東部農林事務所 2017. 鳥取県東部地区の林業の概要. 6pp.
Ueda H, Takatsuki S, Takahashi Y. 2002. Bark stripping of hinoki cypress by sika deer in relation to snow cover and food availability on Mt. Takahara, central Japan. Ecol Res 17:545– 551
Ueda H, Takatsuki S, Takahashi Y. 2003. Seasonal change in browsing by sika deer on hinoki cypress trees on Mount Takahara, central Japan. Ecol Res 18:355–364
Waller DM and Alberson WS, 1997, The white-tailed deer: a keystone herbivore. Wildl Soc Bull 25:217-26.
人工林が広がる場所でのシカの食性 – 鳥取県東部の事例 --
高槻成紀(麻布大学いのちの博物館)・永松 大(鳥取大学)
摘要:日本各地でシカが増加し、森林植生に強い影響を与えるとともに林業被害も増加している。スギ人工林が卓越する鳥取県東部の若桜町では過去20年前からシカが急増し、林床植生が貧弱化した。林業被害対策にはメカニズム解析が不可欠で、シカの食性は一つのポイントとなるが、人工林の卓越する場所でのシカの食性は知られていない。植生は、スギ人工林では柵外はコバノイシカグマ以外は非常に乏しかったが、柵内にはチヂミザサ、ススキ、スゲ類などがあった。落葉広葉樹林でも貧弱で、ムラサキシキブなどが散見される程度であったが、柵内ではタケニグサ、ベニバナボロギク、ジュウモンジシダ、ガクウツギ、ニシノホンモンジスゲ、ススキなどがやや多かった。シカの糞分析の結果、シカの糞組成は植物の生育期でも緑葉が20-30%程度しか含まれておらず、繊維や枯葉の占有率が大きいことがわかった。夏に葉の占有率がこれほど小さいのは神奈川県丹沢のシカで知られているだけである。
はじめに
シカ類の増加と森林生態系の問題は世界的に深刻である(McShea and Rappole, 1992, Waller and Alberson, 1997, Côté et al., 2004, Tanentzap et al., 2010)。日本でも過去20年ほどでシカが増加し(++)、森林生態系に利用を及ぼすとともに(Takatsuki, 2009)、林業の被害を起こしている(大井 1999、Akashi N, Nakashizuka T (1999), Ueda et al., 2002, 2003, 尾崎 2004、Honda et al., 2008)。シカ(ニホンジカ)は落葉広葉樹林に生息することが多いが、場所によってはスギやヒノキの人工林が卓越した場所がある。人工林は密植されるため林内が暗く、林床植物は乏しいことが多い。そのため同じ生息密度であっても広葉樹林よりはシカの食物は乏しく、そのことが林業被害とも関連している。林業被害は被害発生のメカニズムを解明することが重要であり(Ueda et a., 2002, 2003, Honda et al., 2008)、シカの密度や食性を解明は重要な情報となる。
シカの食性は北海道から屋久島まで広範に分析され、大体の傾向は把握されているが(Takatsuki, 2009)、まだまだ残された地域も多い。中国地方はその一つで、2000年に山口県のシカで断片的な情報が報告されたにすぎない(Jayasekara and Takatsuki, 2000)。この分析がなされた1990年代後半には中国地方でのシカの生息は限定的であったが、その後、徐々に拡大した。鳥取県においても兵庫県から連続的な分布域が県東部から徐々に拡大傾向にある(鳥取県, 2017)。
このため農林業への被害が大きくなり、鳥取県ではその抑制のために捕獲が進められ、2010年からは3000頭台、2013年以降は4000頭を超えるレベルになっている(鳥取県 2017)。
調査地である若桜町を含む鳥取県東南部で群落調査とシカの糞密度の関係を調査したところ、若桜町はその中でもシカ密度が高く、植物への影響も強いことが示された(川嶋・永松, 2016)。場所によってはもともとはササがあったが、シカによって食べ尽くされ、低木層も貧弱化し、不嗜好植物(シカが嫌って食べない植物)が増えている場所もあった。
シカが高密度になり、植生に強い影響を与えて食物が乏しくなった状況で死体の胃内容物を調べた例では、枯葉が大きな割合を占めていた(Takahashi and Kaji, 2001)。このようにシカは食物供給の状態に応じて食性を変えることができる。人工林が卓越する場所では食物供給状態は良くないと想定され、シカの食性も落葉樹林とは違う可能性がある。しかし、これまで人工林が卓越した場所でのシカの食性は知られていない。
本調査はこのような背景から鳥取県東部の若桜町のシカの現時点での食性を明らかにすることを目的とした。
方法
1)調査地と林床植生
調査地は鳥取県東部の若桜町に選んだ。この地方は伝統的に林業が盛んであり、若桜町の林野率は95%,人工林率は58%に達していて(鳥取県東部農林事務所 2017),「若桜の杉の美林」として知られる。鳥取県東部のスギは常緑であり葉の垂直的厚さがあるために林床は暗く、林床植生は貧弱である。シカの糞は若桜町役場の南側にそびえる鶴尾山(452 m)の山頂周辺,国指定史跡「若桜鬼ヶ城」の隣接地で採集した(図1)。
図1. シカ糞採取地の位置
糞採取した場所はアカマツとコナラの林で、下生えは強いシカの影響を受けて貧弱になっており、場所によっては2m程度のシカの口が届く高さ以下では植物が非常に乏しくなって「ディア・ライン」が認められた(図2)。
図2. シカ糞採集地の景観。下生えは非常に貧弱で、「ディア・ライン」がみられる場所もある。
シカによって影響を受けている林床群落を記述するために、スギ人工林と落葉広葉樹林の林床に1m四方の方形区をランダムに10個とり、方形区内の植物種の被度(%)と高さを記録した。また史跡保護のために設置されている防鹿柵を利用して,比較のために柵設置後10年の柵内でも同様の記録をとった。これらをもとに、バイオマス指数(被度%×高さcm, Takatsuki and Sato 2013)を求めた。そしてバイオマス指数を木本、双子葉草本、グラミノイド(イネ科とカヤツリグサ科の総称)、シダ、つるに分け、草本はさらに大型草本(50cm以上になるもの)と小型草本に分けて比較した。
2)糞分析
シカの糞の採取に際しては1回分の排泄と判断される糞塊から10粒を採集して1サンプルとし、10サンプルを集めた。糞は2018年の5月から2019年1月まで6回採集した。5月は春、6月、7月、9月が夏、11月が秋、1月が冬に対応する。これを光学顕微鏡でポイント枠法(Stewart, 1967++)で分析し、占有率を求めた。ポイント数は200以上とした。
糞中の成分は次の14群とした。
ササの葉、イネ科の葉、スゲの葉、単子葉植物の葉、双子葉植物の葉、常緑広葉樹の葉、枯葉、その他の葉(コケ、シダなど)、果実、種子、その他、木質繊維、稈、不明
枯葉は黒褐色の不透明な葉脈となった落葉樹の葉であり、緑葉は葉肉部もあり、葉脈は半透明であるから区別ができた。中間的なものもあったが、違いが不明瞭なものは緑葉にした。占有率が1回でも10%以上になった食物群を「主要食物」とし、月間の占有率を多重比較した。
結果
1) 林床植生
調査地のスギ人工林と落葉広葉樹林のようすを図3に示した。柵外はどちらの林でも植物が非常に乏しかった。柵内では回復が見られ、スギ人工林ではススキが、落葉広葉樹林では草本類やスゲが目立った。
図3. 林床の景観。A. スギ人工林、B. スギ人工林柵内、C. 落葉広葉樹林、D. 落葉広葉樹林の柵内
スギ人工林と落葉広葉樹林で植被率(%)を見ると、シカの影響を受けている柵外ではスギ林が3.8%、落葉樹林が3.0%といずれも小さく、両者に有意差はなかった(多重比較、クラスカル・ウォリス検定、z = 0.495, P = 0.96)。柵の内外ではいずれも有意差があった(スギ林、z = 3.631, P = 0.002、広葉樹林、z = 3.9621, P < 0.001)。
スギ人工林と落葉広葉樹林で、バイオマス指数を生育型ごとに合計したところ、スギ人工林でも落葉広葉樹林でも柵外では植物が非常に乏しいこと、柵内では大幅に増加することがわかった(図4)。スギ人工林柵外でのバイオマス指数は45.6で柵内の11.7%に過ぎなかった。内訳ではコバノイシカグマが68.3%を占め、目立って多かった。柵内ではスギ人工林ではイネ科が目立って多く、シダ、大型草本がこれに次いだ。
落葉広葉樹林でのバイオマス指数は13.0と少なく、柵内のわずか2.5%に過ぎなかった。内訳ではムラサキシキブなどの木本類が半量程度であった。柵内では大型草本、シダ、木本が多かった。
柵内をスギ林と落葉広葉樹林で比較すると、スギ林でグラミノイド(チヂミザサ、ススキ、ニシノホンモンジスゲ)が多いこと、広葉樹林で大型草本(タケニグサ、ベニバナボログクなど)、木本(ガクウツギ、クロモジなど)、シダ(ジュウモンジシダ)が多い傾向があった。つる植物は柵外にはほとんどなかったが、柵内ではバイオマス指数で10から20程度あった。
図4. スギ林(Con)と落葉広葉樹林(Bro)の柵外と柵内のバイオマス指数
2)シカの糞の組成
主要食物(一度でも10%以上になった食物)にはイネ科の葉、双子葉植物の葉、枯葉、稈、木質繊維の5つであった。その季節変化は以下の通りであり(図5)、主要種を含む各食物群の組成は付表1に示した。
イネ科は季節変化が不明瞭で10%前後を推移したが、9月の3.8%は7月の10.3%より有意に少なかった(多重比較、z=3.403, p=0.009)。
双子葉植物は7月から11月に多い山型を示した(図3)。6月(1.7%)から7月(15.6%)には有意に増加(z=-3.781, p=0.002)、7月(15.6%)から9月(10.7%)に有意に減少(z=3.326, p=0.011)、9月(10.7%)から11月(19.5%)に有意に増加(z=-3.780, p=0.002)、11月(19.5%)から1月(4.6%)に有意に減少(z=3.78, p=0.002)と、増減を繰り返した。
枯葉は春(5月, 17.8%)と夏(9月、25.1%)、秋(11月、19.8%)に20%前後と多かった。しかし隣あう採集月で有意差はなく、有意差があったのは6月(10.0%)と9月(25.1%)の間だけであった(z=-2.948, p=0.038)。
稈は全体に大きな値をとり、夏に多くなる山型をとった(図3)。7月には45.4%と非常に多く、9月(23.4%)に有意に少なくなり(z=3.780, p=0.002)、1月(10.8%)は11月(27.2%)より有意に少なかった(z=3.024, p=0.030)。
木質繊維も全体に多く、特に5月(55.5%)と1月(48.4%)に多くてU字型を示した。6月(36.3%)から7月(4.2%)へは有意に減少(z=3.781, p=0.002)したが、9月(30.9%)には有意に増加し(z=-3.781, p=0.002)、11月(36.3%)には減少(z=3.780, p=0.002)、1月(36.3%)には増加(z=-3.780, p=0.002)と変化した。
図5. 若桜町(鳥取県東部)のシカの主要食物の月変化
考察
群落調査の結果、柵外ではスギ林でも落葉広葉樹林でも植被率が3%程度しかなく、シカの食物という点で言えば、ほとんど食物がないといえる状況であった。特に面積的に広いスギ林ではその乏しい植物のバイオマス指数の半量以上がコガノイシカグマで占められていた。コバノイシカグマはシカが食べず、食べ残された状況にあった。関東地方ではオオバノイノモトソウが同位的な位置にある(高槻、未発表)。
柵内はスギ林でも広葉樹林でも林床のバイオマス指数が大幅に増加し、スギ林では8.6倍に、広葉樹林では37.9倍になった。このこともシカの採食圧の強さを示している。柵内で回復した植物の構成には違いがあったが、これは元々の種組成の違いと、柵設置後の光条件が落葉広葉樹の方が良いことなどが関係していると考えられる。
シカの糞組成において、夏を中心に葉が多く、冬を中心に繊維が多いというパターンは各地のシカの食性で見られるものであった。しかし葉が夏でも30-40%しかなく、9月には16.8%に過ぎなかった点(後述)と、繊維が冬や春には50%前後にもなり、9月でも30.9%になった点は注目に値する。さらに、夏でも枯葉が食べられて、7月には12.9%、9月には25.1%にも達した。これらのことは、本調査地のシカの食物供給状態が劣悪であることを示唆する。
このことを確認するために、岩手県の五葉山(Takatsuki, 1986)、山梨県の乙女高原(Takahashi et al., 2013)、宮城県の金華山(Takatsuki, 1980)、神奈川県の丹沢山地(高槻・梶谷、印刷中)を比較する。五葉山と乙女高原はシカの密度はあまり高くなくて植物が豊富な場所である。金華山はシカの密度が50頭/km2にも達し、植物は強い影響を受けてシカの食物供給は劣悪である。丹沢山地は場所によって密度は違うが場所によっては金華山並みの高密度の場所もあり、シカの影響を長く受けているので植生は貧弱である(村上ほか, 2007)。
葉の占有率は五葉山が一年を通じて非常に多いが、これはミヤコザサが主体であった(図6)。乙女高原では夏・秋が40%程度、春・冬が50-60%で冬の方がササを食べてやや多くなった。金華山では夏・秋に50-60%と多く、春・冬は20-40%であった。丹沢山地では春と夏が20%程度と少なく、秋・冬にササを食べて40-50%になった。これらに比べると。若桜では春は丹沢と金華山よりは多く、乙女高原程度であり、少ないとは言えなかった。しかし夏は丹沢とともに他の3カ所よりもはるかに少なかった(13.0%)。秋には増加したが5カ所中最少であった(36.3%)。冬も5カ所中最少であった(19.3%)。
図6. 若桜以外4カ所のシカ糞における葉(A)と繊維(B)の占有率。ただし金華山では繊維のデータがない。
繊維は金華山では糞組成を類型するとき「その他」にまとめたので図示できなかった。それ以外の4カ所の比較では、シカの密度が低い五葉山と乙女高原では一年中10%以下と少なかったが、丹沢山地では春以外は多く、若桜では春と冬は4カ所中で最多、夏と秋は丹沢山地に次いだ。
川嶋・永松(2016)は若桜を含むこの地域の植生を調べて、シカによってササが減った場所があると指摘した。シカの糞におけるササの占有率は最も多かった冬でも2.1%に過ぎなかった。しかも、サンプル間のばらつきが大きく、変動係数は繊維が22%であったのに対して、ササは119%であった。このことは調査地ではササが点在していたことと符合する。これは五葉山や乙女高原でササ(ミヤコザサ)が林床全面を覆うのと違い、供給が確実、安定的でないことを意味する。ササは常緑であり、鹿の冬の重要な食物である。このことは五葉山(Takatsuki, 1986)、日光(Takatsuki, 1983)、大台ケ原(Yokoyama et al., 1996)などで示されている。ササの喪失は調査地のシカにとっても林業被害にとっても深刻であろう。
冬は草本類が枯れ、落葉樹は葉を落とすからシカの食物が乏しくなる。そのような状況では植林木への被害が強くなる(Ueda et al. 2002)。この意味で、ササの減少は林業被害につながる可能性がある。
林床群落は非常に貧弱であり、シカの採食影響が強いことを示していた。このことは当然、シカの食性に影響を与えているはずであり、夏でも葉が少なく、枯葉が多いこと、繊維含有率が高いことなどはこのことを反映していると考えられる。
シカの個体数管理は密度を調べて過剰と判断されれば捕獲頭数を決めて駆除がおこなわれる。しかし、シカにとっての資源量である植物量が違えば、密度のもつ意味が違い、植物量の少ない人工林では同じ頭数のシカがいても食物量は少なく、被害が発生する可能性が大きい。その意味でシカ頭数管理は単純に密度ではなく、シカの栄養状態、繁殖率、食性など、シカの状態の質的判断に基づくべきである。この調査の食性分析は、この地方のシカの食料状況が劣悪であることを示した。糞分析方は非侵襲的で比較的簡単であるから応用されることが期待される。
文献
Akashi N, Nakashizuka T (1999) Effects of bark-stripping by sika deer (Cervus nippon) on population dynamics of a mixed forest in Japan. For Ecol Manage 113:75–82
Jayasekara, P. and S. Takatsuki. 2000. Seasonal food habits of a sika deer population in the warm temperate forest of the westernmost part of Honshu, Japan. Ecological Research, 15: 153-157.
川嶋淳史・永松 大. 2016. 鳥取県東部におけるシカの採食による植生の被害状況. 山陰自然史研究, 12: 9-17.
McShea, W. J. and J. H. Rappole. 1997. Herbivores and the ecology of forest understory birds. In The Science of Overabundance: Deer Ecology and Population Management, ed. W. J. McShea, H. B. Underwood and J. H. Rappole. Washington: Smithsonian Institution Press, pp. 268-309.
Stewart, D. R. M. 1967. Analysis of plant epidermis in faeces: a technique for studying the food preferences of grazing herbivores. Journal of Applied Ecology 4: 83–111.
Takahashi,H. and K. Kaji. 2001. Fallen leaves and unpalatable plants as alternative foods for sika deer under food limitation. Ecological Research, 16: 257-262.
Takahashi K, Uehara A, and Takatsuki S. 2013. Food habits of sika deer at Otome Highland, Yamanashi, with reference to Sasa nipponica. Mammal Study, 38: 231-234
Takatsuki, S. 1980. Food habits of Sika deer on Kinkazan Island. Science Report of Tohoku University, Series IV (Biology), 38(1): 7-31.
Takatsuki S. 1986. Food habits of Sika deer on Mt. Goyo. Ecological Research, 1:119-128
Takatsuki, S. 2009. Geographical variations in food habits of sika deer: the northern grazer vs. the southern browser. Sika Deer: Biology and Management of Native and Introduced Populations, (eds. D. R. McCullough, S. Takatsuki and K. Kaji): 231-237. Springer, Tokyo. Elsevier
Takatsuki, S. and T. Kajitani. in press, Food habits of sika deer of Mt. Tanzawa: a case study at a habitat affected by long-term heavy deer grazing. ++ in press
Takatsuki, S. and M. Sato. 2013. Biomass index for the steppe plants of northern Mongolia. Mammal Study, 38: 131-133.
鳥取県. 2017. 鳥取県特定鳥獣(ニホンジカ)管理計画.
鳥取県東部農林事務所 2017. 鳥取県東部地区の林業の概要. 6pp.
Ueda H, Takatsuki S, Takahashi Y. 2002. Bark stripping of hinoki cypress by sika deer in relation to snow cover and food availability on Mt. Takahara, central Japan. Ecol Res 17:545– 551
Ueda H, Takatsuki S, Takahashi Y. 2003. Seasonal change in browsing by sika deer on hinoki cypress trees on Mount Takahara, central Japan. Ecol Res 18:355–364
Waller DM and Alberson WS, 1997, The white-tailed deer: a keystone herbivore. Wildl Soc Bull 25:217-26.