








奈良公園の飛火野と春日山林内のシカの食性
高槻成紀・前迫ゆり
奈良公園のシカの食性については、古く1978年に報告した(高槻・朝日 1978)。これはニホンジカの食性の定量的分析の最も古い論文の一つである。そこで明らかになったのは、奈良公園の平坦地ではシバを主体とした食物組成であること、若草山では春にススキが多くなるということであった。奈良公園のシカは少なくとも数百年あいだ、奈良の人々に手厚く守られることで、公園のシバなどのイネ科を主要な植物としてきた。そして、非常に高密度で生息している。このことは、人と大型野生動物との共存という意味で注目に値することである。同時に, このことは難しい問題を発生してもいる。奈良公園の背後には春日山がある。春日山の森はこの地方の原生的な植生であり、価値が高いが、高密度の奈良のシカが侵入して、森林植生に強い影響を及ぼし、森林の構造だけでなく、維持という点でも問題がある。照葉樹林の更新過程で、シカが食べないナギやナンキンハゼが定着し、場所によっては置き換えが起きている(前迫 2022)。ところが春日山に生息するシカの食性は全く知られていない。
春日山の保全について長年調査してこられた前迫氏との共同で、春日山のシカの食性調査をすることになり、2023年5月から季節ごとにシカのフンを回収して分析したので、その結果を報告する。
調査地
調査地は飛火野と春日山の2カ所とした。調査地2は春日山の西側標高300mのツクバネガシ・コジイ林である(図1)。

図1. シカの糞採集地
飛火野は平坦地でシバ群落が優占し(図2A)、観光客が多く、シカ密度も高い。調査地2は山腹斜面でコジイなどのブナ科常緑樹が卓越し、下層植生としてはイヌガシ、シキミ、アセビなどが生育し、草本層は貧弱であった(図2B)。

図2. 調査地の景観. A: 調査地1、B: 調査地2
方法
調査地で新鮮なシカの糞を探し、糞1群から10粒を10群から採集した。これを0.5 mm間隔のフルイ上で水洗し、植物片を光学顕微鏡で検鏡し、ポイント枠法で評価した。ポイント数は100以上とした。食物はシバ、その他のイネ科、その他のグラミノイド、その他の単子葉類、常緑広葉樹、その他の双子葉植物、針葉樹、枯葉、果実、種子、繊維、稈、不明、不透過に分けた。「不明」は葉や果実ではなく、光学顕微鏡で見て透過性があるもので、特定できないもので、破砕した種子や芽鱗の断片なども含む可能性がある。「不透過」は真っ黒に見えるもので、ドングリの種皮などを含む可能性がある。
またシカの食物供給量の指標として、調査地の地表群落の記述をした。各地に1 m四方の方形区をランダムに10個とり、出現種の(%)と高さ(cm)を記録した。この被度と高さの咳をバイオマス指数(高槻 2009)とて平均値を算出した。そして生育型(沼田 1969)を修正して種ごとにまとめた。
またシカの利用度の指標としてシカの糞粒密度を測定した。調査地1では2 m四方の方形区、調査地2では糞粒が少ないので10 m四方の方形区を5つとり、糞粒の平均値を求めた。この調査は2023年7月から1月までおこなった。
結果
1.シカの糞組成
調査地1(飛火野)ではシバが30%以上を占めたことが最も特徴的だった。特に5月と7月には40%以上となった。そのほかでは5月に彼は、繊維が多かった。7月にはシバ以外では稈が多くなった。10月になるとシバがやや減少し、稈が32.2%と大幅に増え、シバ以外のイネ科も13.2%を占めた。しかし1月になるとシバは大幅に減少し(3.6%)、繊維が39.2%を占めたほか、不明も27.0%と大きく増えた。シバは生産性が高いが、現存量は小さいため、冬になって枯れるとシカの食物としての価値を失い、シカはおそらく木本植物の枝などを食べるために繊維が増えるものと考えられる。

図3. 調査地1(飛火野)のシカの糞組成
一方、調査地2(春日山)では調査地1とは大きく違い、シバはほとんど検出されなかった。5月に多かったのは、繊維(47.7%%)と常緑広葉樹(17.8%)で、シカは林内で常緑樹の葉を枝とともに採食することが多いと考えられる。7月になるとイネ科や落葉樹などが増えるものと想定していたが、全く違い、繊維が60.%もの大きい割合を占め、枯葉も11.8%であり、シカの食糧事情は非常に劣悪であることがわかった。10月になっても緑葉は増えず、繊維は減少したものの不明物が28.9%に増加した。また不透過物も31.0%と多くなった。この不透過物はドングリの殻(種皮)などを含む可能性がある。1月も同様であったが、不明がさらに増えた(43.7%)。

図4. 調査地2(春日山)のシカの糞組成
以上の結果から、調査地1、2での季節間の変化の指標としてWhittakerの類似度指数(PS)を求めたところ、調査地1では春から秋まではPSは80%近かったが、冬になると23.8%と大きく落ち込んだ(図5)。これに対して調査地2では春と夏は71.3%%であったが夏から秋で46.3%に減った後、冬には81.5%に大きく上昇した。

図5. 調査地1(飛火野)と調査地2(春日山)における季節間の糞組成の類似度指数
一方、調査地1と調査地2との類似度を季節ごとに見ると、春から秋までは30%前後と違いが大きかったが、冬には69.6%と大きくなった。これは春から秋までは調査地1ではシバが主要であり、調査地2では樹木の繊維や葉、不明物が多く、組成の違いが大きかったが、冬にはどちらでも繊維が多くなったことを反映しているためである。

図5. 調査地1(飛火野)と調査地2(春日山)における糞組成の類似度指数の季節変化
2.供給量
調査地1でのバイオマス指数の季節変化は春から秋までは300-500と非常に大きかったが、冬になるとシバが蹴れたため激減した(図6)。
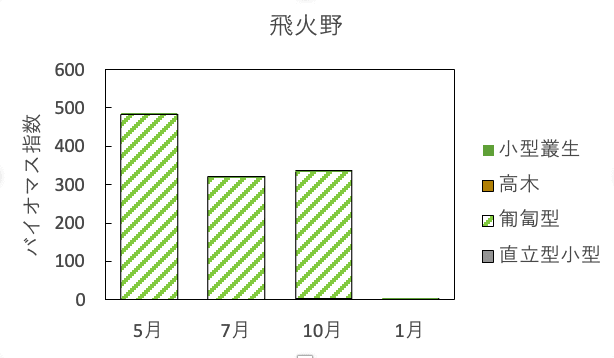
図6. 調査地1(飛火野)におけるバイオマス指数の季節変化
一方、調査地2では春は高木と低木が少量あるだけだが、夏になるとシダと香木が増えてピークとなった。ただし値は調査地1の半量以下であった。その後、秋には減少し、冬には常緑高木がわずかになった(図7)。

図7. 調査地2(春日山)におけるバイオマス指数の季節変化
3.糞粒密度
調査地1では10月には460/4m2と非常に多く、11月に半分程度に減少してそのまま維持された(図8)。一方、調査地2では10/4m2以下と非常に少なかったが、1月には490/4m2ほどと非常に多くなり、調査地1を上回った(ただし、この時は時間の関係で1か所しか調べていないので、今後追加調査をする予定)。つまり、春日山の林内では夏秋はシカがあまり利用しないが、冬になると大きく利用度が高くなることがわかった。
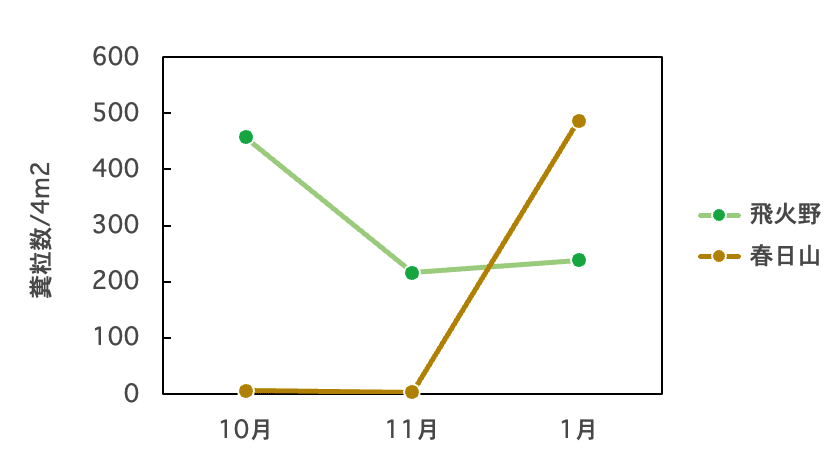
図8. 調査地1(飛火野)と調査地2(春日山)における糞粒密度の季節変化(暫定)
考察
<飛火野のシカの食性>
飛火野では春と夏にはシバが40%を前後を占めたが、半世紀ほど前の1976年5月の糞組成でもシバが春は30-40%、夏には70-80%占めていた(高槻・朝日 1978)。シバは生産力が高く(吉良 1952, Inoue et al. 1975, 高槻 2023)、シカは生育期にはシカにとって非常に重要な食物となりうる。しかし、現存量が小さいため、枯れると飼料価値は大幅に減る。実際、糞組成でも冬にシバは激減したし、バイオマス指数の変化もこれを裏付けていた。宮城県金華山においては、夏にシバ群落を利用していたシカが冬には周辺の森林を利用するようになることが知られている(Ito and Takatsuki 2005)。調査地1でも夏から秋まではシカの糞粒密度が高く、それは1月まで維持された。この点は金華山と違うが、飛火野ではシバが枯れても、観光客がシカ煎餅を与えるなどの事情でシカがシバ群落にとどまるものと考えられる。
<春日山のシカの食性>
春日山のシカの食性は知られていなかったが、今回初めて明らかになった。それによると、春日山のシカの糞組成は飛火野のそれとは全く違い、イネ科はほとんど出現せず、5月に常緑広葉樹が10-18%を占めた。他の季節では葉は少なく、繊維や不明物が多かった。これらは概ねバイオマス指数と対応したが、夏にはバイオマス指数がある程度大きくなったにもかかわらず、糞組成には葉は少なかった。その理由は林床では木本実生やシダが多くなったが、これらはシキミ、イヌガシ、イワヒメワラビ、コバノイシカグマなど不嗜好植物であり、シカは食べない。このためバイオマス指数が大きくてもシカの食物としては価値がない。
調査地2は若草山と500 mほどしか離れていない。若草山にはススキなどシカの食物になる植物が多く、実際シカも多くいる。調査地2と若草山の距離はシカにとって移動は十分可能であると思われるが、糞組成にはイネ科はほとんど検出されなかったことから、春日山のシカは行動上の理由から自由に行き来をするということはないようである。
<シカによる春日山の利用の季節変化>
春日山の森林は冬でも食物が増えるわけではないが、シカの利用度が著しく高くなった。これはシカが林内で寒さを防ぐとか林床の果実などを探して食べるためなどの理由でシカ利用度が非常に高くなるものと考えられる。
<シカによる春日山の森林に及ぼす影響>
この森林は林内が暗いため、シカが採食する植物が乏しく、シカは常緑樹などを探して食べるが、量的には少ない。その時に枝先も同時に摂取するため、糞中に繊維が多くなるものと思われる。春、夏は林床にある落ち葉も採食するため、糞中に枯葉が5-15%程度検出されたが、秋冬は枯葉も5%未満になった。
<飛火野のシカと春日山のシカの関係>
この分析により、春日山林内のシカの食性は奈良公園の平坦地のものとはまったく違い、シカは林内にとどまって乏しい植物を食べていることがわかった。もし、シカが林外で植物を食べて林内では休息や睡眠だけをするのであれば、後述する森林への影響は問題とならないが、実際にはシバ群落にいるシカはおもにシバを、森林にいるシカは基本的に森林の植物を食べていた。このことは、春日山の原生的常緑広葉樹の更新のためには問題がある。常緑広葉樹林は林床が暗く、採食を受けた植物の回復力が乏しい。同じ常緑広葉樹林である宮崎県椎葉にある九州大学の演習林ではシカが増加してからスズタケが壊滅的な影響を受けたことが知られているが(猿木ほか 2004)、これも回復力の弱さが関係している。春日山の林では時間をかけて更新する過程で後継樹が供給できず、場所によってシカの不嗜好植物であるナギやナンキンハゼに置き換わることが知られている(前迫 2022)。そのため樹冠を形成するシイ、カシなどの幼樹が生育できず、健全な森林更新ができず、一種の偏向遷移が起きているといえる、
<奈良のシカと春日山森林の保全>
奈良のシカは長い時間をかけて形成された人とシカとの共存の例として広く知られ、観光的にも価値があり、その保護が必要であろう。しかし、同時に春日山原生林も世界遺産としての価値がある。奈良公園の自然を管理する上では、この両方を実現することに難しさがある。その両立のためには、現状を把握し、何が起きているかの客観的把握が必要である。本調査により、春日山のシカの行動権の把握も解明すべき課題であることが示された。もし森林の更新が必要であると判断されたのであれば、ある程度広いシカ排除柵を設置することなどにより、後継樹の確保をするなどの対策が必要になるだろう。
文献
Inoue, Y., M. Iwamoto, T. Kaminaga and S. Ogawa. 1975. Measurement and comparison of primary production of semi-natural pastures dominated by Zoysia japonica. In Ecological Studies in Japanese Grasslands, with Special Reference to the IBP Area (ed. Numata, M.), JIBP Synthesis, Vol. 13: 123-124.
Ito, T. Y. and S. Takatsuki. 2005. Relationship between a high density of sika deer and productivity of the short-grass (Zoysia japonica) community: a case study on Kinkazan Island, northern Japan. Ecological Research, 20: 573-579. DOI:10.1007/s11284-005-0073-6
吉良龍夫. 1952. 生態学的に見たいわゆる過放牧牧野. 小物生態学報, 1: 209-213.
前迫ゆり. 2022. 照葉樹林に侵入した外来木本種の拡散にニホンジカが与える影響. 日本生態学会誌, 72: 5-12.
沼田真(編). 1967.『図説植物生態学』, 朝倉書店
猿木重文・井上晋・椎葉康喜・長澤久視・大崎繁・久保田勝義. 2004. 九州大学宮崎演習林においてキュウシュウジカの摂食被害を受けたスズタケ群落分布と生育状況 2003年調査結果. 九州大学演習林報告, 85: 47-57.
高槻成紀. 2009. 野生動物生息地の植物量的評価のためのバイオマス指数について. 麻布大学雑誌,19/20: 1-4. https://agriknowledge.affrc.go.jp/RN/2030800112.pdf
高槻成紀. 2022. ススキとシバの摘葉に対する反応−シカ生息地の群落変化の説明のために. 植生学会誌, 39: 85-91. https://www.jstage.jst.go.jp/article/vegsci/39/2/39_85/_article/-char/ja
高槻成紀・朝日稔.1978. 糞分析による奈良公園のシカの食性,II.季節変化と特異性. 「天然記念物「奈良のシカ」報告(昭和52年度」:25-37.








※コメント投稿者のブログIDはブログ作成者のみに通知されます