








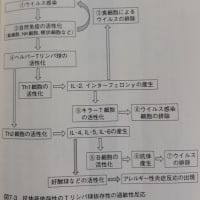
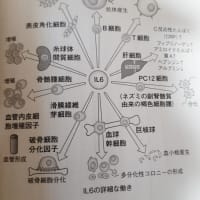
獲得免疫は脊椎動物が持つ「特異的生体防御システム」です。獲得免疫の主役はリンパ球のB細胞とT細胞であり、それぞれ液性免疫応答と細胞性免疫応答と呼ばれています。液性免疫応答は、B細胞が作る「抗体」(血液中に分泌され体内を循環する)により抗原を認識して攻撃します。細胞性免疫応答は、細胞傷害性T細胞(キラーT細胞)が感染された又は変性した自己細胞を認識することによりその自己細胞を攻撃します。またヘルパーT細胞はキラーT細胞やB細胞の制御の役割をします。
特異的生体防御システムの特徴は以下のようです。
(1)特異性
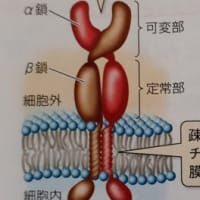
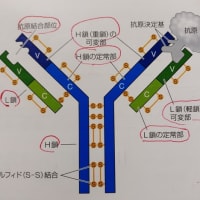
T細胞受容体とB細胞受容体及び抗体は、抗原(病原体などの標識)を個別具体的に認識します。その方法は、抗原が例えばタンパク質であれば、多数存在するアミノ酸の数個の配列の局所領域を認識するものです。このような認識する局所領域を「抗原決定基」(エピトープ)といいます。抗原のタンパク質には複数の抗原決定基があることがあり、その中でも特に強く免疫応答を示す部分のものを「主要抗原決定基」といいます。
(2)自己と非自己の識別
獲得免疫システムは、ヒトの体内に何万個もの自己抗原である異なったタンパク質もあるため、自己と非自己(異性化した自己含め)を正確に認識して、自己抗原を攻撃しないような厳格なシステムがあります。胸腺での未熟T細胞の選別(自己抗原を攻撃しないような)では、作られた多種類のT細胞のうち生き残れるのは2、3%とされています。
(3)多様性
T細胞受容体とB細胞受容体(抗体)は、様々な非自己抗原である病原体、異物、ウイルス、細菌などの抗原決定基を個別具体的に認識するために、遺伝子組換え(VDJ組換え)や抗体遺伝子への変異導入(AID 活性化誘導シチジンデアミナーゼ )により、無限ともいえる(通説では10億種類)非自己抗原に適合できるような多種の細胞が用意されています。そのため、どのような非自己抗原が侵入してきても、その抗原決定基に適合するT細胞受容体やB細胞受容体(抗体)が準備されています。
(4)免疫記憶
ある病原体が最初に侵入すると、それに適合したT細胞やB細胞が活性化(細胞増殖)されますが、攻撃用の細胞(短期間(数日)でなくなる)の他に、記憶用の細胞も作られてゆっくり分裂を継続します(数十年以上)。その記憶細胞があるため、二度目にその病原体が侵入したときは、即時に免疫システムが発動するようになります。
") | ゲノムが語る生命像 (ブルーバックス) 本庶 佑 講談社 |
「獲得免疫系の第一の目的は、無限に存在するかもしれない外来微生物等の抗原を、いかにきちんと認識するのかという基本的な仕組みを確立することである。このためには、抗原受容体の種類がきわめて多種類存在することが必要である。限られた遺伝情報の中から、どうやって何千万、何億もの種類の抗原受容体を生み出すことができるのであろうか。この謎に対する答えとしては、遺伝子の断片の組み合わせによって新たな遺伝子を生み出すという巧妙な方法が、利根川進らによって明らかにされた。
「VDJ組換え」と呼ばれるこの遺伝子組換えの仕組みは、抗体遺伝子(B細胞受容体)とT細胞受容体遺伝子の両方に用いられており、この組換えを行うRAG1とRAG2という酵素もBリンパ球とTリンパ球に共通である。
このような遺伝子組換えが起こる仕組みは、脊椎動物のかなり早い時期に始まったと思われる。しかし、脊椎動物の祖先系と考えられる脊椎動物である
ヤツメウナギやメクラウナギなどには、この仕組みは存在しない。またRAG1とRGA2の遺伝子にはイントロンがなく、遺伝子の組換えを行う仕組みが決まったDNA配列を目印とした断片のつなぎ合わせ様式となっている。これらの特徴が、トランスポゾンと呼ばれる自ら遺伝子改変を行う仕組みときわめて似ていることから、その進化の原点はトランスポゾンが脊椎動物の祖先系に侵入したことに起源があるのではないかと考えられている。つまり、今日地球上に存在する脊椎動物の祖先のどこかで、このようなトランスポゾンの感染が生殖細胞に起こり、今日地球上に存在する脊椎動物はすべてその子孫であるという驚くべき推論であるが、今日それが多くの研究者の支持を受けている。
…抗体を作るBリンパ球の中では、抗原刺激によってAID(活性化誘導性シチジンデアミナーゼ)という分子の発現が誘導される。AIDの発現により、抗体遺伝子に2つの遺伝的な変異が導入される。第一は、抗体の可変部、すなわち抗原認識部位に塩基の置換(体細胞突然変異)が導入され、抗原と結合力の高い抗体を作り出す。先のVDJ組換えによって作り出された抗体レパートリーの中から、抗原を認識した細胞がAIDを発現し、その細胞の抗体遺伝子に、さらに細かい塩基の変異が導入される。
…AIDの発現により第二の変化は、抗体のクラスを変えることである。通常のBリンパ球は免疫ブログリン(Ig)のうちIgMを産生しているが、抗原刺激によってIgG、IgE、IgAなどの抗体を産生する細胞に変わる。この変化は「クラススイッチ」と呼ばれ、この際には遺伝子の大幅な欠失を伴うDNAの組換えが起こる。このようなクラスの変化(クラススイッチ)は、結合した抗原をどのような仕組みで処理するかという、抗体の効果の多様性を生み出す。また体の特定な場所に特化した機能を生み出す。
…AIDの誕生は、RAG1、RAG2より進化の上で古いことが明らかになった。すなわち、脊索動物であるヤツメウナギやメクラウナギにすでにAIDの祖先型が存在している。興味深いことに、これらの生物における抗原受容体は、RAG1、RAG2によって作られる今日型の抗原受容体とまったく異なる構造をし、非常に強い接着性を持ったVLRと呼ばれる分子であることがクーパーらによって明らかにされた。VLRは、AID祖先型分子により遺伝子断片の情報をつなぎ合わせる遺伝子変換と呼ばれる遺伝子再構成を用いながら抗原受容体を作り上げていたのである。
この仕組みは、今日、AID自身の働きにも引き継がれており、体細胞突然変異や遺伝子変換、クラススイッチにおける遺伝子切断などの仕組みは、おそらく基本的に同じものであったと考えられる。
…VLRの消失の理由は、先に述べたRAG1、RAG2を含むトランスポゾンの感染が脊椎動物の初期段階で起こったことにより、新しい免疫受容体の多様化機構が生じたことによると思われる。(引用終わり)」
「VDJ組換え」と呼ばれるこの遺伝子組換えの仕組みは、抗体遺伝子(B細胞受容体)とT細胞受容体遺伝子の両方に用いられており、この組換えを行うRAG1とRAG2という酵素もBリンパ球とTリンパ球に共通である。
このような遺伝子組換えが起こる仕組みは、脊椎動物のかなり早い時期に始まったと思われる。しかし、脊椎動物の祖先系と考えられる脊椎動物である
ヤツメウナギやメクラウナギなどには、この仕組みは存在しない。またRAG1とRGA2の遺伝子にはイントロンがなく、遺伝子の組換えを行う仕組みが決まったDNA配列を目印とした断片のつなぎ合わせ様式となっている。これらの特徴が、トランスポゾンと呼ばれる自ら遺伝子改変を行う仕組みときわめて似ていることから、その進化の原点はトランスポゾンが脊椎動物の祖先系に侵入したことに起源があるのではないかと考えられている。つまり、今日地球上に存在する脊椎動物の祖先のどこかで、このようなトランスポゾンの感染が生殖細胞に起こり、今日地球上に存在する脊椎動物はすべてその子孫であるという驚くべき推論であるが、今日それが多くの研究者の支持を受けている。
…抗体を作るBリンパ球の中では、抗原刺激によってAID(活性化誘導性シチジンデアミナーゼ)という分子の発現が誘導される。AIDの発現により、抗体遺伝子に2つの遺伝的な変異が導入される。第一は、抗体の可変部、すなわち抗原認識部位に塩基の置換(体細胞突然変異)が導入され、抗原と結合力の高い抗体を作り出す。先のVDJ組換えによって作り出された抗体レパートリーの中から、抗原を認識した細胞がAIDを発現し、その細胞の抗体遺伝子に、さらに細かい塩基の変異が導入される。
…AIDの発現により第二の変化は、抗体のクラスを変えることである。通常のBリンパ球は免疫ブログリン(Ig)のうちIgMを産生しているが、抗原刺激によってIgG、IgE、IgAなどの抗体を産生する細胞に変わる。この変化は「クラススイッチ」と呼ばれ、この際には遺伝子の大幅な欠失を伴うDNAの組換えが起こる。このようなクラスの変化(クラススイッチ)は、結合した抗原をどのような仕組みで処理するかという、抗体の効果の多様性を生み出す。また体の特定な場所に特化した機能を生み出す。
…AIDの誕生は、RAG1、RAG2より進化の上で古いことが明らかになった。すなわち、脊索動物であるヤツメウナギやメクラウナギにすでにAIDの祖先型が存在している。興味深いことに、これらの生物における抗原受容体は、RAG1、RAG2によって作られる今日型の抗原受容体とまったく異なる構造をし、非常に強い接着性を持ったVLRと呼ばれる分子であることがクーパーらによって明らかにされた。VLRは、AID祖先型分子により遺伝子断片の情報をつなぎ合わせる遺伝子変換と呼ばれる遺伝子再構成を用いながら抗原受容体を作り上げていたのである。
この仕組みは、今日、AID自身の働きにも引き継がれており、体細胞突然変異や遺伝子変換、クラススイッチにおける遺伝子切断などの仕組みは、おそらく基本的に同じものであったと考えられる。
…VLRの消失の理由は、先に述べたRAG1、RAG2を含むトランスポゾンの感染が脊椎動物の初期段階で起こったことにより、新しい免疫受容体の多様化機構が生じたことによると思われる。(引用終わり)」
「V(D)J遺伝子再構成(英:V(D)J recombinationまたはsomatic recombination)は、免疫システム内の免疫グロブリン(Ig)・TCR(T細胞受容体)生成の初期ステージにおける遺伝子再構成の仕組み。初期のリンパ組織(骨髄ではB細胞、胸腺ではT細胞)で起こる。
V(D)J遺伝子再構成は、骨髄や胸腺でのリンパ球の遺伝子断片(V、D、J)のランダムな組み合わせである。抗体やTCRをコードするDNAは多数の断片として染色体の上に並んでいる。これらのDNA断片はリンパ球の分化の過程で連結され、完全なDNAとなる。いろんな遺伝子をランダムに選べるので、いろんなタンパク質をつくり、いろんな抗原(バクテリア、ウイルス、寄生菌、腫瘍、花粉など)に対抗することができる。」
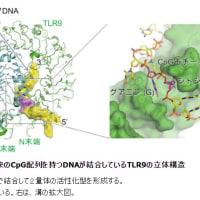
「活性化誘導シチジンデアミナーゼ(かっせいかゆうどうシチジンデアミナーゼ、Activation-Induced (Cytidine) Deaminase、AID)は、DNA中のシチジン基からアミノ基を取り除く(脱アミノ)、24 kDaの酵素である。
AIDは現在、二次抗体多様化のマスター制御因子であると考えられている。AIDがその開始に関与しているのは、3つに分かれた免疫グロブリン(Ig)多様化プロセス、体細胞超変異(SHM)、クラススイッチ組換え(CSR)、遺伝子変換(GC)である。」
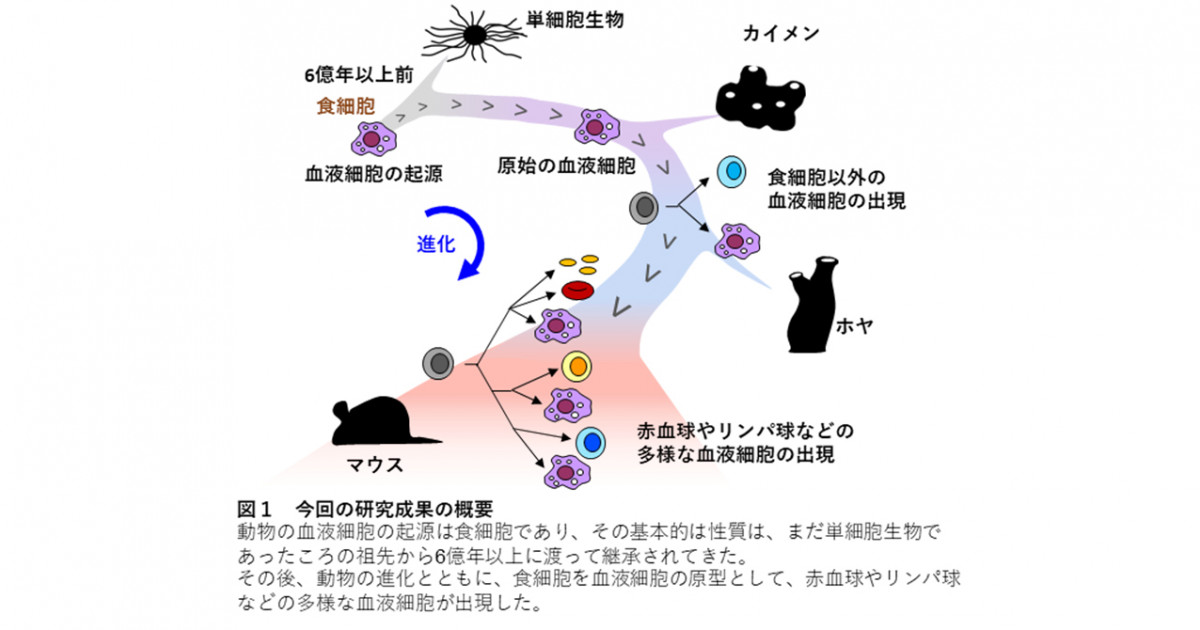
第16話_いろいろな生物の免疫の仕組み
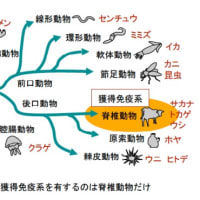
「まず動物のおおまかな分類を解説する。 構造的に、口がないもの(海綿動物)、口が肛門を兼ねるもの(腔腸動物)、口と肛門があるものに分けられ、口と肛門のある ものの中では発生過程で口が先にできる のが前口動物で、後にできるのが後口動物 である(図 1)。
全ての動物は自然免疫系を有するが、獲得免疫系を有するのは脊椎動物だけで、種の数としては動物界全体の数%にすぎな い。ほとんどの無脊椎動物は自然免疫系だ けで生きているのである。
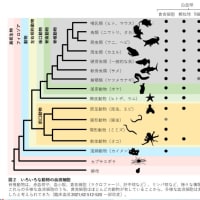
…軟骨魚類以上の脊椎動物は、すべて同じ タイプの獲得免疫系を有している。種による違いをみていこう。全ての種において T 細胞は胸腺でつくられており、T 細胞系は 大枠ではほぼ同じである。
…一方 B 細胞のつくられ方は、種による違 いが大きい。…マウス やヒトでは
遺伝子断片がランダムに組み 換えられる様式で多様性がつくられる。しかしニワトリでは多様性は組み換えでは 生じず、遺伝子変換と体細胞超変異でつくられる。前者は一度できた抗体遺伝子の一部分を後で入れ替えるという様式で、後者は点突然変異を誘導するという様式である。哺乳類でもウサギ、ウシのように主に遺伝子変換と体細胞超変異で多様性を得る種もいる。つくられる場所も、マウスや ヒトでは骨髄だが、ニワトリではファブリキウス嚢という肛門の近くの器官、ウサギ では虫垂、ヒツジでは腸管のパイエル板と、 大きく異なっている。







※コメント投稿者のブログIDはブログ作成者のみに通知されます