








1.インターフェロン
情報伝達物質(サイトカイン)の1種であるインターフェロンは、1954年に日本の長野泰一氏と小島保彦氏が「ウイルス干渉因子」として発見されました。
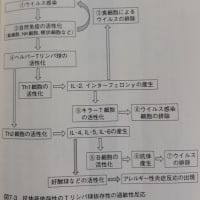
ウイルス感染により、Toll様受容体(TLR)のTLR3、TLR7、TLR9(エンドソームに存在)やRIG-I、MDA-5らのパータン認識型の受容体が情報を感知すると、I型インターフェロンが活性化されます。また様々な抗原が侵入したときに産生されるIL-1、IL-2、IL-12、TNF、CSFなどのサイトカインによっても、インターフェロンは誘導されます。
(1)IFN typeⅠ (Ⅰ型インターフェロン)
I型インターフェロンには、インターフェロンα(IFN-α)、インターフェロンβ(IFN-β)などがあります。
一般的にインターフェロンというと、このⅠ型インターフェロンのことをいいます。Ⅰ型インターフェロンには主に3つの機能があります。
①直接ウイルス複製を抑制する。
●RNaseL(リボヌクレアーゼ L)の活性化により、ウイルスのmRNAを
分解する。
●真核生物蛋白質合成開始因子 eIF-2α(eIF2S1)をリン酸化してウイルス
ペプチド鎖の合成開始を阻止する。
●その他、MxAが誘導されて、ウイルス感染細胞のアポトーシスの促進や
ウイルス増殖の抑制が行われます。
②NK細胞がウイルス非感染細胞を攻撃しないように、MHCクラスI分子
の発現を増加させる。
③NK細胞を活性化させてウイルス感染細胞を除去する
(2)IFN typeⅡ(2型インターフェロン)
II型インターフェロン(IFN-γ)は、免疫系の細胞によって分泌されて、マクロファージを活性化します。
2.ウイルスによるインターフェロン抑制
ウイルスにはインターフェロンを抑制する機能を持つものがいるようです。
インターフェロンの抑制方法としては、(1)IFN 結合性蛋白の産生、(2) JAK/STAT 系に関わる蛋白の分解、(3)JAK/STAT 系に関わる蛋白の活性化阻止、(4)活性化転写因子の核内移行の阻害、(5)JAK/STAT 系のネガテブ制御因子の誘導などがあるようです。
スペイン風邪を復元して実験した結果では、インターフェロンが抑制されていたことが分かりました。
また新型コロナウイルスも、インターフェロンを抑制する機能を持っているようです。
")
「…新型コロナウイルス感染ではⅠ型インターフェロンがうまく作られないことがあるようです。…ウイルスが作るタンパク質の一つであるORF3bが、宿主細胞のⅠ型インターフェロン遺伝子の活性化を抑えて、Ⅰ型インターフェロンの産出を抑えます。また、同じくウイルス由来の別のタンパク質PLProが、ORF3bとは別のメカニズムで、Ⅰ型インターフェロン遺伝子の活性化を抑え、結課としてⅠ型インターフェロンがうまく作られなくなります。
Ⅰ型インターフェロンは、ウイルス増殖を抑えるだけでなく、周囲の細胞に対して炎症性サイトカイン産生を促してウイルスに対する炎症反応を促進する役目があることから、Ⅰ型インターフェロンが十分にできないと、抗ウイルス反応がうまく起きないだけでなく、風邪症状も起こりにくくなり、その間にウイルスは局所で増えていくことになります。」
「…驚くべきことに、SARS-CoV-2のORF3bタンパク質は、SARS-CoVのORF3bタンパク質よりも強いインターフェロン阻害活性があることを見いだしました。また、コウモリやセンザンコウで同定されている、SARS-CoV-2に近縁なウイルスのORF3bタンパク質についても同様に解析した結果、SARS-CoV-2のORF3bタンパク質と同様、強いインターフェロン阻害活性があることを明らかにしました。」
「ORF3bはサルベコウイルス亜属のコロナウイルスに見られる遺伝子で、短い非構造タンパク質をコードしています。これはSARS-CoV ( SARSという病気を引き起こす) とSARS-CoV-2 ( COVID-19を引き起こす)の両方に存在します …インターフェロンアンタゴニストと同様の効果がある 」
「パパイン様プロテアーゼ(パパインようプロテアーゼ、英: papain-like protease、PLpro)は、コロナウイルスの複製に不可欠なシステインプロテアーゼで、CAグループペプチダーゼ(C16ファミリー)に属する。
…PLproの触媒ドメインの分子構造は、「広げた右手」がN末端で「ユビキチン様」ドメイン(UBL)に結合して構成されている。PLproの構造は、脱ユビキチン化酵素(DUB)の構造と似ている。さらに、PLproの酵素活性のin vitro特性評価によって、このタンパク質様プロテアーゼはユビキチン(Ub)およびUBLのISG15(英語版)(インターフェロン誘導遺伝子15、interferon-induced gene 15)タンパク質を認識して、加水分解することがわかった。
UbとISG15はどちらも、ウイルス感染に対する宿主の自然免疫系の免疫応答のシグナル伝達において重要な役割を果たしており、これらの欠如はウイルス増殖に有利に働くことが分かっている。」
")
「もし私たちがウイルスと呼ぶ病原体が生き物というなら、ウイルスを人工合成した彼らは生き物を創造した「神」になるのだろうか。2005年秋、インフルエンザ・ウイルスを再合成してみせた米陸軍病理研究所のJ・K・タウベンバーガーたちのことである。
スペイン風邪を引き起こしたウイルス…1997年、アラスカから持ち帰った組織片からほぼ完全なウイルスを発見したタウンバーガーは歓喜して、ウイルスの遺伝子を解読し始めた。…すべての解読を終えたのは2005年。解読がどれほど大変だったかは歳月が示している。
タウンバーガーたちは…比較的、簡単にDNA版のウイルス遺伝子を作ることができた。彼らに協力してウイルスの遺伝情報をもとにDNAを作り、これをプラスミドに組み込む重要な仕事をこなしたのは米ニューヨークのマウント・サイナイ医科大学の研究者たち。そのプラスミドを人の腎臓細胞の中に注入するという最後の仕上げ的な作業を担ったのは、世界の感染症対策の総本山と目される米疾病管理センター(CDC)のテレンス・タンビーらだった。
…ここからのプロセスはウイルスが感染した細胞の中で増殖するのと同じだ。
…復元されたウイルスは予想通り、致命的な毒性を持っていた。タンビーたちが、復元したウイルスを実験動物のネズミに感染させたところ、数日後にはネズミの肺に通常のインフルエンザ・ウイルスと比べ数万倍のウイルスが発生。ネズミは肺炎を起こして死んでしまった。
2007年1月…米国のタウンバーガーたちが公表していたウイルスの遺伝情報をもとに、河岡はウイルスを復元、その成果を英科学誌ネイチャーで発表したのだ。
…河岡は二つの点で異彩を放った。人工合成したウイルスを人間に近いカニクイザルに感染させた点と、ウイルスの強力な毒性の背後に免疫の異常反応がからんでいるのを突きとめたことだ。
…まずスペイン風邪ウイルスをサルの鼻などに接種、すると24時間以内に体力や食欲が弱まり、感染して1週間ほどたつと重い肺炎を起こし呼吸困難になった。回復は不能と判断した研究者たちはここで実験を停止、サルを安楽死させることとなった。
解剖するとサルの肺は約7割の領域が肺炎に冒され、水分がいっぱいたまってた。肺にたまった水分でサルが呼吸不全に陥ったことを示唆する光景だった。通常のインフルエンザ・ウイルスを接種したサルが肺炎を起こすこともなく軽い症状にとどまったのと比べると症状は重篤。スペイン風邪ウイルスは通常のウイルスに比べ、気管や肺で百倍以上に増えていた。
…免疫は異常な反応を見せていた。通常、ウイルスなどが侵入すると、生き物の体にはウイルスを抑えるインターフェロンという情報伝達物質が現れる。インターフェロンの別名はウイルス抑制因子。しかしサルの体内ではインターフェロンの分泌が抑制されていた。
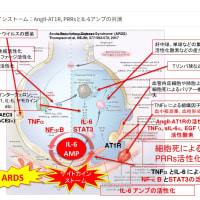
一方、過剰に分泌が増えた情報伝達分子もあった。発熱・腫れ・むくみ・痛みなどを引き起こすインターロイキン6という分子だ。その様子は関係者が嵐に例えて「サイトカイン・ストーム(嵐)」と呼んだほどだった。
…炎症は免疫の働きに不可欠な営みであるが、インターロイキン6がこれほど過剰に分泌され、またウイルスの増殖を抑えるインターフェロンの分泌が抑制されてはサルの体が無事ですむわけもなかった、と考えられる。」
スペイン風邪を引き起こしたウイルス…1997年、アラスカから持ち帰った組織片からほぼ完全なウイルスを発見したタウンバーガーは歓喜して、ウイルスの遺伝子を解読し始めた。…すべての解読を終えたのは2005年。解読がどれほど大変だったかは歳月が示している。
タウンバーガーたちは…比較的、簡単にDNA版のウイルス遺伝子を作ることができた。彼らに協力してウイルスの遺伝情報をもとにDNAを作り、これをプラスミドに組み込む重要な仕事をこなしたのは米ニューヨークのマウント・サイナイ医科大学の研究者たち。そのプラスミドを人の腎臓細胞の中に注入するという最後の仕上げ的な作業を担ったのは、世界の感染症対策の総本山と目される米疾病管理センター(CDC)のテレンス・タンビーらだった。
…ここからのプロセスはウイルスが感染した細胞の中で増殖するのと同じだ。
…復元されたウイルスは予想通り、致命的な毒性を持っていた。タンビーたちが、復元したウイルスを実験動物のネズミに感染させたところ、数日後にはネズミの肺に通常のインフルエンザ・ウイルスと比べ数万倍のウイルスが発生。ネズミは肺炎を起こして死んでしまった。
2007年1月…米国のタウンバーガーたちが公表していたウイルスの遺伝情報をもとに、河岡はウイルスを復元、その成果を英科学誌ネイチャーで発表したのだ。
…河岡は二つの点で異彩を放った。人工合成したウイルスを人間に近いカニクイザルに感染させた点と、ウイルスの強力な毒性の背後に免疫の異常反応がからんでいるのを突きとめたことだ。
…まずスペイン風邪ウイルスをサルの鼻などに接種、すると24時間以内に体力や食欲が弱まり、感染して1週間ほどたつと重い肺炎を起こし呼吸困難になった。回復は不能と判断した研究者たちはここで実験を停止、サルを安楽死させることとなった。
解剖するとサルの肺は約7割の領域が肺炎に冒され、水分がいっぱいたまってた。肺にたまった水分でサルが呼吸不全に陥ったことを示唆する光景だった。通常のインフルエンザ・ウイルスを接種したサルが肺炎を起こすこともなく軽い症状にとどまったのと比べると症状は重篤。スペイン風邪ウイルスは通常のウイルスに比べ、気管や肺で百倍以上に増えていた。
…免疫は異常な反応を見せていた。通常、ウイルスなどが侵入すると、生き物の体にはウイルスを抑えるインターフェロンという情報伝達物質が現れる。インターフェロンの別名はウイルス抑制因子。しかしサルの体内ではインターフェロンの分泌が抑制されていた。
一方、過剰に分泌が増えた情報伝達分子もあった。発熱・腫れ・むくみ・痛みなどを引き起こすインターロイキン6という分子だ。その様子は関係者が嵐に例えて「サイトカイン・ストーム(嵐)」と呼んだほどだった。
…炎症は免疫の働きに不可欠な営みであるが、インターロイキン6がこれほど過剰に分泌され、またウイルスの増殖を抑えるインターフェロンの分泌が抑制されてはサルの体が無事ですむわけもなかった、と考えられる。」
「…コロナウイルスは、ウイルス感染の最初の10日間に自然免疫を回避します。感染の初期段階では、SARS-CoV-2 は、ヒト細胞における弱い IFN-I 誘導物質であるSARS-CoVよりもさらに低いI 型インターフェロン(IFN-I) 応答を誘導します。 SARS-CoV-2 も IFN-III 反応を制限します。加齢に伴う形質細胞様樹状細胞の数の減少は、新型コロナウイルス感染症(COVID-19)の重症度の増加と関連しているが、これはおそらくこれらの細胞が実質的なインターフェロン産生細胞であるためであると考えられる。
生命を脅かす新型コロナウイルス感染症患者の 10 パーセントは、I 型インターフェロンに対する自己抗体を持っています。
IFN-I 応答の遅延は、 COVID-19疾患の後期段階で見られる病原性炎症 (サイトカインストーム)の一因となります。ウイルス感染前 (または感染の非常に初期段階) での IFN-I の適用は、PEG 化 IFN-λIII による治療と同様に防御効果があり、ランダム化臨床試験で検証されるべきである 。」
『2. ウイルスによるインターフェロン情報伝達系 抑制の分子機構』
「ウイルスに対する細胞や生体の防御機構の中で、インターフェロン(IFN)の示す抗ウイルス活性は大変重要であり、またIFNの多面的作用は獲得免疫発動にとっても欠かせないものでもある。ウイルスは IFN の情報伝達系を抑制することによってほぼ全 IFN システムを抑制する機能を獲得してきた と思われる。これまで判明している IFN 情報伝達系の抑制機構は、(1)IFN 結合性蛋白の産生、(2) JAK/STAT 系に関わる蛋白の分解、(3)JAK/STAT 系に関わる蛋白の活性化阻止、(4)活性化転写 因子の核内移行の阻害、(5)JAK/STAT 系のネガテブ制御因子の誘導、に整理される。我々の検討 している HSV1 ではネガテブ制御因子である SOCS3 の誘導が、ムンプスウイルスは V 蛋白による STAT-1,STAT-3 の分解が、麻疹ウイルスでは V 蛋白、C 蛋白が IFN レセプターと複合体を形成し Jak-1 のリン酸化が阻止されていることが、IFN 情報伝達系抑制に関わっている。
…ウイルスや 細菌による IFN 産生制御,IFN による TLR や MyD88 の制 御は,免疫機構が微生物感染により広範な影響を受けるこ とを示している.ウイルスや細菌は,生体が大きなシステ ムとして構築してきた防御機構を様々な側面から撹乱し自 らの増殖性や侵襲性を確保する能力を備えている.このこ とは,ウイルスが単に IFN による抗ウイルス作用を抑制 するというだけではなく,JAK/STAT 情報伝達系の抑制 を介して IFN シグナルやサイトカインシグナル,さらに は TLR シグナルが乱され多くの細胞機能が抑制あるいは 促進されると共に,Th0 リンパ球から Th1 や Th2 への誘導 レベルの乱れ,樹状細胞機能変化,T リンパ球や B リンパ 球の分化・成熟過程の変化,その結果として免疫機構の破 綻に至ることが考えられる.従って,これらの微生物が如 何にして細胞内情報伝達機構を抑制,阻害するかという分 子機構を理解することは,微生物の病原性を解明する一助 になるであろうし,またより有効な治療法に繋がる可能性 を秘めていると思われる. 」
「インターフェロン(英: interferon、略号:IFN)とは、動物体内で病原体(特にウイルス)や腫瘍細胞などの異物の侵入に反応して細胞が分泌する蛋白質のこと。ウイルス増殖の阻止や細胞増殖の抑制、免疫系および炎症の調節などの働きをするサイトカインの一種である。
…1954年に、伝染病研究所所長(当時)の長野泰一と小島保彦が「ウイルス干渉因子」として発見し報告した。1957年には、イギリスのアリック・アイザックス(Alick Isaacs)やスイスのジャン・リンデンマン(Jean Lindenmann)たちもウイルス増殖を非特異的に(抗体ではない)抑制する因子として確認し、ウイルス干渉(Interference)因子という意味で「Interferon(インターフェロン)」と命名した。 1980年頃に、インターフェロンが悪性腫瘍に効果があることが発見され、抗がん剤として発展していった。 蚕[8] やハムスターの体内にヒトの細胞を埋め込んで、その細胞にC型肝炎ウイルスの遺伝子を組み込んだセンダイウイルスを感染させることにより、インターフェロンを産生させるという方法を利用して大量生産が可能になった。
…ウイルスの感染や2本鎖RNAなどによって直接誘導されることが知られている。これらの細胞外での受容体としてはToll様受容体(TLR)でその中でもエンドソームに存在するTLR3、TLR7、TLR9である。また、細胞内に存在する受容体としてはRIG-I、MDA-5が関与し、これらがI型インターフェロンの発現を高めると考えられる。また体内にいろいろな抗原が侵入したときそれに反応してIL-1、IL-2、IL-12、TNF、CSFなどのサイトカインが産生される。インターフェロンの産生はこれらのサイトカインによっても誘導される。
インターフェロンにより調節される細胞内シグナル伝達経路の代表的なものとしてはJAK-STAT経路が知られるが、それ以外の経路も関与していると考えられる。
インターフェロンαとβはリンパ球(T細胞、B細胞)、マクロファージ、線維芽細胞、血管内皮細胞、骨芽細胞など多くのタイプの細胞で産生され特に抗ウイルス応答の重要な要素である(詳しくはI型インターフェロンの項を参照)。インターフェロンαとβはマクロファージとNK細胞をともに刺激し、腫瘍細胞に対しても直接的に増殖抑制作用を示す。
インターフェロンγは活性化されたT細胞で産生され免疫系と炎症反応に対して調節作用を有する。IFN-γにも抗ウイルス作用と抗腫瘍作用があるが弱く、その代わりIFN-αとβの効果を増強する作用がある。IFN-γは腫瘍のある局所で働く必要があり、がん治療への有効性は低い。IFN-γはTh1細胞からも分泌され、白血球を感染局所にリクルートして炎症を強化する作用がある。またマクロファージを刺激して細菌を貪食殺菌させる。Th1細胞から分泌されたIFN-γはTh2反応を調節する作用でも重要である。免疫応答の調節にも関わっており、過剰な産生は自己免疫疾患につながる可能性がある。IFN-ωは白血球からウイルス感染または腫瘍の局所で分泌される。」
「I型インターフェロン(英:type I interferon)とは、インターフェロンファミリーのうち、インターフェロンα(英語版)(IFN-α)とインターフェロンβ(英語版)(IFN-β)などを含めた総称で、ウイルス感染で誘導される抗ウイルス系のサイトカインである。「I型」という名前は、免疫系の細胞によって分泌されマクロファージを活性化するII型インターフェロン(IFN-γ)などと区別するための呼称であるが、一般に「インターフェロン」というとI型インターフェロンのことを指す。
…I型インターフェロンの主な機能としては、
- (1)ウイルス複製を抑制することで、細胞のウイルス抵抗性を上昇させる
- (2)ウイルス非感染細胞のMHCクラスI分子の発現を増加させ、NK細胞の攻撃から保護する
- (3)NK細胞を活性化させてウイルス感染細胞を除去する
という3つである。 まず、I型インターフェロンが細胞に結合すると、(2'-5')オリゴアデニル酸合成酵素系とプロテインキナーゼ系が活性化する。(2'-5')オリゴアデニル酸合成酵素系(2-5AS系)では、通常3'-5'の形で結合しているATPを2'-5'結合オリゴマーに重合させることでエンドヌクレアーゼであるRNaseLを活性化しウイルスのmRNAを分解する。一方プロテインキナーゼ系では、真核生物蛋白質合成開始因子 eIF-2α(eIF2S1)をリン酸化することでウイルスペプチド鎖の合成開始を阻止する。この他にもインターフェロンは抗ウイルス活性を示す遺伝子を誘導する。その遺伝子の1つとしてMxA(myxovirus resistance A)がある。MxAはウイルス感染細胞におけるアポトーシスの促進とウイルス増殖の抑制を促すが、これはMxAが小胞体ストレスを起こすことによるものだと考えられている。
上で述べたような直接的な抗ウイルス活性の他に、I型インターフェロンはウイルス非感染細胞のMHCクラスI分子の発現を高めることでNK細胞から正常細胞を保護している。というのも、NK細胞はウイルスによってMHCの発現が抑制されたり、立体配座(コンフォメーション)を変更させられたMHCを持つ細胞を攻撃する一方で正常のMHCクラスI分子を持っている細胞に対してはNK細胞に抑制性のシグナルが入り攻撃を行わないからである。この一方で、I型インターフェロンはNK細胞を活性化する役割も担っている。ここで活性化されたNK細胞はウイルス感染細胞を除去するとともにインターフェロンγ(IFN-γ)を放出することでT細胞依存性の細胞傷害を誘導する。」
「リボヌクレアーゼ LまたはRNase L (潜在型) は、リボヌクレアーゼ 4または2'-5' オリゴアデニル酸シンテターゼ依存性リボヌクレアーゼとしても知られ、インターフェロン (IFN)誘導性リボヌクレアーゼであり、活性化されると細胞内のすべてのRNAを破壊します(両方とも細胞性そしてバイラル)。RNase L は、ヒトではRNASEL遺伝子によってコードされる酵素です。」
「インターフェロン誘導 GTP 結合タンパク質 Mx1 は、ヒトではMX1遺伝子によってコードされるタンパク質です。
マウスでは、インターフェロン誘導性 Mx タンパク質が、インフルエンザ ウイルス 感染に対する特異的な抗ウイルス状態を担っています。さらに、ヒトオルソログ MxA は、動物由来のインフルエンザウイルスの主要な決定因子です。この遺伝子によってコードされるタンパク質は、その抗原関連性、誘導条件、物理化学的性質、およびアミノ酸分析によって決定されるように、マウスタンパク質に類似している。この細胞質タンパク質は、ダイナミン スーパーファミリーと大型GTPaseファミリーの両方のメンバーです。」
「真核生物翻訳開始因子 2 サブユニット 1 (eIF2α) は、ヒトではEIF2S1遺伝子によってコードされるタンパク質です。
この遺伝子によってコードされるタンパク質は、翻訳開始因子 eIF2 タンパク質複合体のアルファ (α) サブユニットであり、タンパク質合成開始の初期の制御されたステップを触媒し、イニシエーター tRNA (Met-tRNA i Met ) の40S リボソームサブユニットへの結合を促進します。 」
「インターフェロンは感染細胞から分泌され、周囲の細胞に対し自身が感染したことを警告する。また免疫機構の細胞がこれにより刺激されると、ウイルス監視の一環としてインターフェロンを分泌する。インターフェロンは細胞表面にある受容体と結合する小さなタンパク質である。この信号は細胞内へと伝えられ、これによってウイルス防御に関わる何百ものタンパク質が作られる。細胞が作るインターフェロンにはいくつかの種類がある。インターフェロン-α(interferon-alpha、PDBエントリー 1itf)とインターフェロン-β(interferon-beta、PDBエントリー 1au1)は最も一般的な型で、大半の細胞種、特に免疫機構細胞によって作られる。これらは成長を止め防御に集中するための基本的な信号を送る。一方インターフェロン-γ(interferon-gamma、PDBエントリー 1rfb)は主にT細胞から分泌され、免疫機構の反応を調節する信号を送る。
…ウイルスは巧妙で、想像の通り、様々な方法で進化してインターフェロンによる防御に対抗する。ウイルスによってインターフェロンの活動を阻害する段階は異なり、インターフェロンがその受容体に結合するところから、最終的に核に達する一連の信号伝達経路に至るまでさまざまである。例えば、右図に示したタンパク質(PDBエントリー 3bes)はハツカネズミ(mouse)に天然痘(smallpox)に似た症状(マウス痘、奇肢症、ectromeria)を引き起こすウイルス(エクトロメリアウイルス ectromeria virus、ECTV)から得られたものである。このタンパク質(青)はインターフェロン(赤)を捕獲し、受容体へ結合するのを阻害する。 」
『1. 自然免疫による核酸認識』
「ウイルスや細菌のもつ核酸(DNA と RNA)は自然免疫系により認識され,I 型インターフェロン や炎症性サイトカインが産生され,感染病原体に対する生体防御応答が誘導される.我々は発現スク リーニングにより,二重鎖 DNA に対する自然免疫応答を制御する細胞内分子として TRIM56 を同定 した.
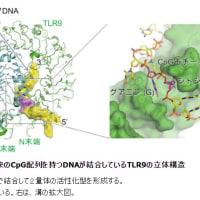
TRIM56 はユビキチンリガーゼとして機能し,STING と呼ばれるアダプター分子の K63 型ユ ビキチン化を促進した.この修飾により,TBK1 キナーゼがリクルートされ最終的に I 型インターフェ ロンが誘導された.以上のことから,DNA に対する自然免疫応答において,TRIM56 によるユビキ チン化を軸とした新たなシグナル伝達経路の存在が明らかとなった.一方,Toll-like receptor (TLR) 7 と TLR9 は,ウイルスの核酸を認識し,プラズマ細胞様樹状細胞から I 型インターフェロン産生をさ せる.我々は,抗ウイルス因子として報告されていた Viperin が,プラズマ細胞様樹状細胞において TLR7/9 を介した I 型インターフェロン産生に重要な役割を果たしていることを見出した.Viperin は, プラズマ細胞様樹状細胞において TLR7/9 の刺激より転写因子 Interferon regulatory factor (IRF) 7 依 存的に強く誘導され,脂肪滴に局在している.Viperin は,プラズマ細胞様樹状細胞において TLR7/9 の下流で働き IRF7 を活性化するシグナル伝達因子として知られている TRAF6 と IRAK1 に 結合し,これらの因子を脂肪滴上へとリクルートする.その結果,IRAK1 の K63 結合型ユビキチン 化が効率的に誘導され,IRAK1 による IRF7 の活性化を介した I 型インターフェロンの産生が促進さ れる.Viperin が,直接的なウイルス複製阻害に加えて,TLR7/9 を介した I 型インターフェロン産 生の促進により抗ウイルス応答に関わっていることが判明した. 」
「…Viperinは直接的なウイルス複製の阻害にくわえ,TLR7およびTLR9を介したI型インターフェロンの産生の促進により抗ウイルス応答にかかわっていることが明らかになった.」
「ラジカル S-アデノシル メチオニン ドメイン含有タンパク質 2 は、ヒトでは RSAD2遺伝子によってコードされるタンパク質です。RSAD2 はウイルスプロセスにおける多機能タンパク質であり、インターフェロン刺激遺伝子です。
…Viperin は、ウイルス RNA 依存性 RNA ポリメラーゼ (RdRp) を阻害するチェーン ターミネーターddhCTP (3'-デオキシ-3',4' ジデヒドロ-CTP) を生成できるラジカル SAM 酵素です。この活性はアミノ酸の代謝とミトコンドリア呼吸を無効にするようです。
インフルエンザウイルスの出芽と放出の阻害において、ビペリンは、イソプレノイド生合成経路に必須の酵素であるファルネシル二リン酸シンターゼ(FPPS)に結合して酵素活性を低下させることにより、細胞の原形質膜上の脂質ラフトを破壊することが示唆されています。[13]ビペリンは、宿主タンパク質 hVAP-33 およびNS5A との相互作用および複製複合体の形成の破壊を介して、HCV のウイルス複製を阻害することが示唆されています。」
「JAK-STATシグナル伝達経路 (ジャック-スタット・シグナルでんたつけいろ)は細胞外からの化学シグナルを、細胞核に伝え、DNAの 転写と発現を起こす情報伝達系。 免疫、増殖, 分化、アポトーシス 、発癌などに関与する。 JAK-STATシグナルカスケードは主に3つの構成要素からなる: 細胞表面の受容体、 Janus kinase (JAK)、2つの信号トランスデューサおよび転写活性化(STAT)タンパク質である。[1] JAK-STAT機能が損なわれたり、制御できないと、自己免疫疾患, 免疫不全症候群や悪性腫瘍などを引き起こされることがある。 」
「MyD88(myeloid differentiation factor 88)はTLRやIL-1ファミリーサイトカイン受容体の下流でシグナルを伝えるアダプタータンパク質である。
…ヒトのパターン認識受容体にはTLR、NLR、RLR、CLRなどが知られ、TLRは10個あまり知られている。TLRのうちTLR3とTLR4の一部以外はMyD88を介して転写因子NF-κBを活性化させる。TLR3はMyD88ではなくTRIFという別のアダプタータンパク質を介してⅠ型インターフェロンの産出に関わる。
MyD88を介したシグナルはマクロファージ等からの炎症性サイトカインの産出を誘導し自然免疫応答を惹起するのに加え、樹状細胞の活性化を促し獲得免疫応答を誘導するにも重要な役割を果たす[9]。T細胞依存性抗原に対する抗体産出におけるMyD88の役割は樹状細胞の活性化による獲得免疫応答の誘導ならびにB細胞の活性化による抗体産出の増強がある。
抗原提示細胞のMyD88シグナルの活性化は抗原提示細胞によるサイトカイン産出や補助刺激分子の発現を増加させることで、抗原特異的T細胞の活性化を誘導する。さらにTLR等を介したB細胞のMyD88シグナルの活性化はB細胞とT細胞との会合を促し、またB細胞の胚中心B細胞ならびに形質細胞への分化を促進することが知られている。このようにMyD88シグナルは自然免疫応答惹起だけではなく、獲得免疫応答ならびに抗体産生を正に制御している。」
「NF-κB(エヌエフ・カッパー・ビー、核内因子κB、nuclear factor-kappa B)は転写因子として働くタンパク質複合体である。NF-κBは1986年にノーベル生理学医学賞受賞者であるデビッド・ボルティモアらにより発見された。免疫グロブリンκ鎖遺伝子のエンハンサー領域に結合するタンパク質として発見され、当初はB細胞に特異的なものと考えられていたが、後に動物のほとんど全ての細胞に発現していることが明らかとなった。高等生物に限らずショウジョウバエやウニなどの無脊椎動物の細胞においてもNF-κBが発現している。
NF-κBはストレスやサイトカイン、紫外線等の刺激により活性化される[1]。NF-κBは免疫反応において中心的役割を果たす転写因子の一つであり、急性および慢性炎症反応や細胞増殖、アポトーシスなどの数多くの生理現象に関与している。NF-κB活性制御の不良はクローン病や関節リウマチなどの炎症性疾患をはじめとし、癌や敗血症性ショックなどの原因となり、特に悪性腫瘍では多くの場合NF-κBの恒常的活性化が認められる。さらにNF-κBはサイトメガロウイルス (CMV) やヒト免疫不全ウイルス (HIV) の増殖にも関与している。」
「転写因子(てんしゃいんし)はDNAに特異的に結合するタンパク質の一群である。DNA上のプロモーター領域に、基本転写因子と呼ばれるものと、RNAポリメラーゼ(RNA合成酵素)が結合し、転写が開始する。DNAの遺伝情報をRNAに転写する過程を促進、あるいは逆に抑制する。転写因子はこの機能を単独で、または他のタンパク質と複合体を形成することによって実行する。ヒトのゲノム上には、転写因子をコードする遺伝子がおよそ1,800前後存在するとの推定がなされている。」







※コメント投稿者のブログIDはブログ作成者のみに通知されます