









→ from Columba livia:01
・顎 jaws:
鳥の顎は"歯を欠く"のが特徴的である。
もっとも、胚発生においては、上皮に歯を発生させる能力が残っていることが知られており、
例えばマウスの神経堤細胞を移植することで容易に主竜類型の歯を誘導することができる。
ちなみに中新世にはオステオドントルニス(Osteodontornis; 骨質歯鳥類)なんてのもいたが、
あれは明らかに歯ではなく嘴の縁が二次的に鋸歯状になったものだ。
さて一般に顎の開閉は、単純に1つの蝶番を動かすように、単純で一方向な動きだと思われがちだが、
実際には各動物群ごとで複雑かつ多様な挙動を示す。
顎口類の中でも比較的原始形質を多く留めるとされる軟骨魚でさえ、
複数の関節を介したなかなかに合理的で素敵な顎の動きをする。
知る限り、トカゲなどは顎の開閉のスピードによって真逆の動きをする筋肉まである。
そもそも、我々哺乳類の頭がほぼ全てがガッチリ癒合したものであるので、いまいち想像しがたいのかも知れないが、
大抵の脊椎動物で頭蓋骨というのは関節を持っていたりするものなのである。
鳥の頭蓋は中でも特殊化が進んでおり、
脳幹(braincase)・口蓋と方形骨(palate & quadrate)・上顎(upper jaw)・下顎(mandible)の4つの部位に分かれ、
それぞれが関節(hinge)で接続する。

上図、橙色の丸が関節。赤矢印が筋肉である。
それぞれのパーツが効率よく動き、嘴でよく獲物を捕えて飲み込むことができるようになっている。
・前肢 forelimb:
鳥の骨格の中でも最も独特なのが前肢。
脊椎動物は前肢を交互に、前後に動かして運動するが、
鳥は左右の肢が連動し、しかも運動方向は背腹に限定されている。
これこそ正に、ほかの獣脚類に見られない"鳥的"な特徴であり、飛行には不可欠な形質でもある。
この前足の運動を根元で支えるのが、
上腕骨(humerus)-胸骨(pectoral)を繋ぐ表層の胸筋(Pectoralis)と
深層にある烏口上筋(Supracoracoideus; 要は"笹身")だ。
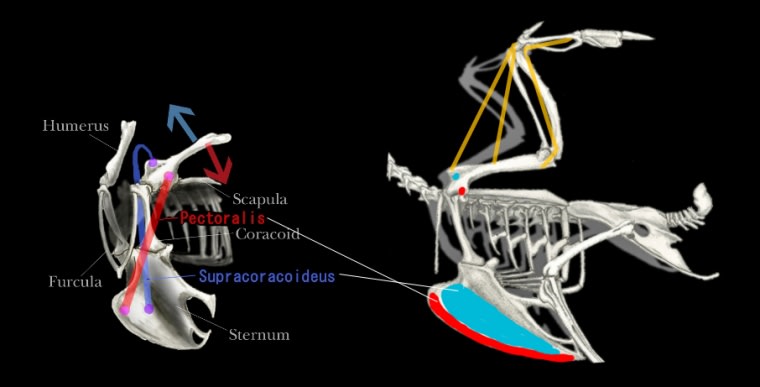
上図で赤色が胸筋、青色が烏口上筋。図の右側の、赤色と青色はそのおよその付着点だ。
矢印は各色に対応した筋肉が収縮したとき、上腕骨がどちらに動くかを示す。
胸筋(Pectoralis)は言わずもがな、
烏口上筋(Supracoracoideus)は、真骨類で胸鰭の屈筋(flexor)から分かれ、四足動物ならば全てが有している筋肉である。
ところが、多くの動物で烏口上筋は上腕骨を体の前(斜め下)方向へ動かす機能を持っているのに対し、
恐らく鳥だけが、背側方向へ打ち出す運動を担っている。
これは筋肉の付着部位や運動方向そのものが特に変化したのではなく、
上腕骨-烏口骨の関節の変化と、
烏口上筋が上腕骨に接続する腱と烏口骨との僅かな位置的なズレに起因するところが大きい。
多くの四足動物でこの腱は烏口骨の外側を真っ直ぐ走るのに対し、
鳥では一部が烏口骨の下(現生鳥類ではここがほぼ孔状になっている)に入り込んでしまっている。
そしてこの烏口骨の構造が、腱に対してちょうど滑車のような役目を果たすため、
烏口上筋により、上腕骨は実際、上側から引っ張られる力を受けることになる。
これは形態上非常に僅かな変化なのだが、運動機能においては非常に重大な変化をもたらしている。
恐らく遺伝子レヴェルでの変化など見つけようにも、まず存在しないぐらいのものであり、
こうした事例がある限りは、分子進化のみならず、形態進化を仔細に老い続ける意義は大きい。
ちなみに上腕骨-胸骨を繋ぐ烏口上筋は、哺乳類に関してはその位置がダイナミックにずれ、
上腕骨-肩甲骨の背を繋ぐ筋肉になっており、
棘上筋(Supraspinatus)・棘下筋(Infraspinatus)の2つに分かれて存在している。
・後肢 hindlimb:
主竜類は、現在知られている限りでは、史上初めて直立2足歩行を行ったグループである。
二足歩行を行う上での利点というのはイマイチ思いつかないが(前足がフリーになるから色々使えるぞというのは二次産物)
何となくだが運動性は高いのかもしれない。
にしてもバランスを保つのは大変なはずだ。
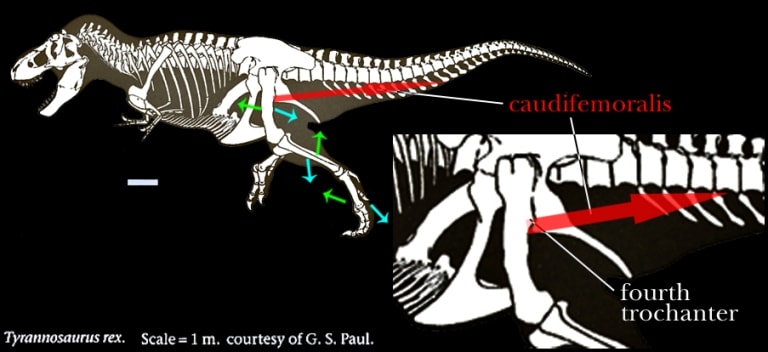
主竜類(Archosauria)の大腿骨(femur)には、第四転子(fourth trochanter)と呼ばれる突起がある。
主竜類と言うからにはワニにも恐竜(特に鳥脚類は顕著な鉤状をしている)にもあって、
ここには尾-大腿骨筋(caudi-femoralis)が付く。
caudifemoralisは要するに尻尾と後脚を繋ぐ筋肉であり、収縮すると大腿骨が後側に引かれる。
下図、ティラノサウルス・レックス(Tyrannosaurus rex)でこれを示した。
収縮したときには、脚の各骨は青色の矢印方向に動く。
ところが、鳥では尾が極端に短縮し、
それと共にcaudifemoralisも矮小化してしまっている。
さらにそれと共に第四転子も消失し、鳥の大腿骨は二次的に平坦な状態になっている。
尾が短縮し、筋肉が縮小することで後脚の運動も大きく変わった。大腿骨をほとんど動かさなくなったのである。
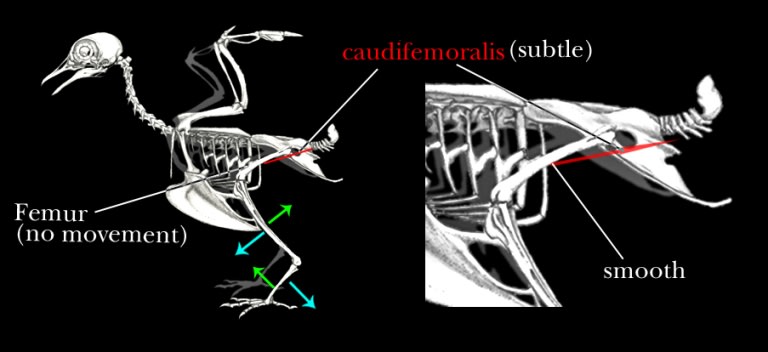
このように、単に尾が短くなることだけで歩行が大きく変わってきてしまうのである。
前脚にせよ後脚にせよ、
これらはある意味、恐竜-鳥を分ける重要なポイントに見えて、実際たぶんそうなのだろうが、
"恐竜"と呼ばれる中にも例えばオヴィラプトル類などはこれらの特徴をバッチリ持っていて、
かと思えば始祖鳥はそうでもなかったりする。
形態の進化も、難しいけれど面白いでしょ?
BLOG内LINK:
・specimens: その他の収蔵標本。
・vertebrata:01・02 脊椎動物。
・disosaurs of gondwana 2009: 『大恐竜展 知られざる南半球の支配者』
・dinosaur expo 2009:01 - 05 『恐竜2009 砂漠の奇跡』
・avian digit: 鳥の指パラドックス。
・gecko:skelett トッケイヤモリ全身骨格
= National Geographic:Today's Photo =
References:
・『Functional Anatomy of Vertebrates: An Evolutionary Perspective 3rd. edition』 (Hardcover)
Warren F. Walker :著/編, Karel F. Liem :編 (Brooks/Cole Pub Co, 2000)
・『The Vertebrate Body』 (Hardcover)
Alfred Sherwood Romer, Thomas S. Parsons :著 (W.B.Saunders, 1977)
・『Studies on the Structure & Development of Vertebrates』(Hardcover)
Edwin S. Goodrich :著 (Macmillan and co., limited, 1930)
・『恐竜学 進化と絶滅の謎』
David E.Fastovsky / David B.Weishampel:著, John Sibbick:イラスト, 真鍋 真:訳 (丸善, 2006)
・『恐竜博2005』パンフレット
・『Developmental Biology eighth edition』 (Hardcover)
Scott F. Gilbert:著, (Sinauer Associates Inc; 8 Har/Cdr版, 2006)










goo.gl/bgwWhC