








●獲得形質の遺伝
周知のようにラマルクは、生物にはその器官を使用するかしないかに応じて変異し、しかもそうして獲得した変異を子孫に伝える能力が備わっている、と主張しました。今日においても、ラマルクの思想の流れを汲む学説は一定数の生物学者に支持されています。この学説によれば、新しい種を生み出す変異は、胚そのものに内在する偶発的なものでもなければ、一定の形質を、有用性とは無関係に一定の方向に発達させる一種独特な決定論に従うものでもありません。それは、生物が自分に課せられた生存条件に適応しようとする努力そのものから生まれます。無論この努力は、外的環境の圧力によって機械的に惹起される或る器官の機械的作用に過ぎない場合もあるでしょうが、意識と意志を含む場合もあるでしょう。この学説を代表する人物の一人であるアメリカの博物学者コープは、努力を後者の意味に解しています。したがって新ラマルク主義そのものは必ずしも内的で心理的な発達原理に訴える必然性は持たないとしても、この学説は現在の進化思想のうち、そうした発達原理を受け入れ得る唯一の学説と言えます。また新ラマルク主義は、それぞれ独立した複数の発達系統において、同一の複雑な器官が形成されることを説明できそうな唯一の学説でもあります。事実、同じ環境を利用しようとする同じ努力が、同じ結果に達するのは自然なことです。さらに外的環境の提出する問題が唯一つの解答しか許さないような性質のものであれば、同じ努力が同じ結果に達する蓋然性はより一層高くなる、と言えるでしょう。とは言え検討すべき問題が一つ残されています。それは、この「努力」という言葉には、どんな新ラマルク主義者が考えているよりももっと深い心理的な意味を賦与しなければならないのではないか、ということです。
実際、単なる大きさの変異と、形態の変化とは区別して考えなければなりません。或る器官の機能を働かせ続ければ、それが強化され大きさを増すことを否定する人はいないでしょう。しかしそのことと、例えば軟体動物の眼や脊椎動物の眼が漸進的に発達することとは全く別のことです。眼の発達の原因を受動的に受け取られた光の影響に求めるならば、たった今批判したばかりの説に逆戻りすることになります。逆に内的作用を持ち出すのであれば、わたしたちが通常努力と呼んでいるものとは全く別のものを問題にする必要があります。何故なら繊毛虫類の眼点から脊椎動物の眼に移行するまでの間には、驚くほど整然とした無数の複雑さの段階が存在するにもかかわらず、努力によって或る器官がほんのわずかでも複雑になったなどという話は聞いたことがないからです。動物に関する限り、進化過程を努力によるものと看做すそうした考え方を認めてもよい知れません。しかしこの考え方を、どうして植物界にまで拡げることができるでしょうか。植物界では、形態の変異は常に機能の変化を含んでいるようには見えませんし、それを伴っているようにも思えません。変異の原因を心理的な性質のものと仮定するのはよいとしても、その原因を努力と呼ぶためには、言葉の意味を異常に拡張しなければならないでしょう。わたしたちがすべきことはこの努力そのものを掘り下げ、もっと深い原因を探ることなのです。
特に規則的に遺伝する変異が問題となる場合には、そういった探究が一段と重要になってきます。わたしたちは、ここで獲得形質の遺伝に関する論争の詳細に立ち入るつもりはありません。わたしたちの専門外の問題について、余り明確な態度を表明するのは本来なら差し控えたいところです。とは言っても、この問題に全く無関心でいるわけにもいきません。今日、哲学者は漠然とした一般論で満足しているわけにはいかないこと、哲学者も科学者の行った実験の細部を辿り、その結果について科学者と意見を交換する義務があること、このことを最も痛感するのが、他でもない、獲得形質の遺伝に関する問題を扱うときです。仮にスペンサーが獲得形質の遺伝に関する問題を最初に自らに課していたとすれば、恐らく彼の進化論は全く違ったものになっていたに違いありません。個体が身に付けた習慣が、極めて例外的な場合にしか子孫に遺伝しない(わたしたちにはそう思えるのですが)ならば、スペンサーの心理学は全面的に修正されなければならないでしょうし、彼の築いた哲学の大部分はその根拠を失ってしまう筈です。そういうわけで、獲得形質の遺伝に関する問題がどのように提起されているか、またこの問題の解決をどの方向に求めるべきかについて、わたしたちの見解を率直に述べて置くことにします。
獲得形質が遺伝するという説はかつて独断的に肯定されていましたが、その後、生殖細胞に備わっているとされる性質からア・プリオリに引き出された理由によって、同じく独断的に否定されるに至りました。ヴァイスマンが、生殖質は連続するという仮説に基づき、生殖細胞――卵と精子――が体細胞からほとんど独立していると結論するに至ったのは周知の通りですが、そこから獲得形質は遺伝しないという主張が広がっていったのです。この主張は、今日においても最も有力な説の一つと言って差し支えありません。――しかし獲得形質が遺伝し得るものであることが万に一つでも実験によって明らかになった場合、生殖質は現在考えられているほど周囲の体細胞から独立していないということになり、事実上、獲得形質は遺伝すると考えてもよい、ということになるでしょう。つまり獲得形質は遺伝するか否かという机上の議論には意味がなく、問題の解決はひとえに実験にかかっている、ということになります。とは言え口で言うのは簡単ですが、難しいのはここから先の部分です。いわゆる獲得形質とは、多くの場合、習慣もしくは習慣の結果を指しますが、獲得された習慣の根底に、自然的な資質なり傾向が存在していないことは滅多にありません。そのため遺伝したのは個体の体(たい。原語はsomaで、生殖細胞を除く体全体)が獲得した習慣なのか、或いは寧ろ、獲得された習慣に先立つ自然的な傾向ではないのか、という疑問が常に付いて回ります。そうした傾向は、既に個体に内属していたのかも知れず、したがってもともとその胚に、そして個体の生殖細胞に内属していたのかも知れません。それゆえモグラの視力が悪いのは、地下で生活する習慣を身に付けたからだ、と結論するのは早計です。恐らくモグラは、眼が衰退しつつあったからこそ地下での生活を余儀なくされたのでしょう。そうだとすれば、視力が失われていく傾向は生殖細胞から生殖細胞へと遺伝したのであって、モグラの体(ソーマ)そのものは何も獲得せず、何も失わなかったと考えるのが自然です。フェンシングの名手の息子が父親よりずっと早く優れた選手になったからと言って、親の習慣が子に遺伝したと決め付けることはできません。というのもその場合、発達途上にある何らかの自然的な傾向が、父親を生み出した生殖質から息子を生み出した生殖質へと遺伝する過程で、起源にあるエランの作用によって成長した結果、言ってみれば息子は父親の身に付けた習慣など見向きもせず父親を凌駕する反射神経を獲得したのかも知れないからです。長期間にわたって行われる動物の家畜化についても、同様のことが言えます。家畜化を目的として、動物の中から或る特定の種やその種の成員の一部を選ぶ際、選定の目安となるのはそれらの持つ自然的性向ですが、遺伝するのはそれらが身に付けた習慣なのか、それとも選定の目安となった或る種の自然的性向なのかを見極めるのは困難です。実を言うと、獲得された特性が遺伝する事例のうち、疑わしい事例や、様々な解釈を容れる余地のある事実をすべて除外すると、絶対に異論を挟む余地のないものとして残るのは、何人もの生理学者によって繰り返し再現され、確認されているブラウン・セカールの有名な実験くらいしかありません。彼の実験が明らかにしたところによれば、モルモットの脊髄もしくは坐骨神経を切断すると癲癇症状が現れ、子孫にもこの症状が遺伝します。坐骨神経や索状体などの損傷は、モルモットに様々な障害を惹き起こしましたが、これらの障害は、時として最初の障害とは異なる形、例えば眼球突出や足指喪失といった形で子孫に遺伝することもあったと言います。――しかしブラウン・セカールが提示したこれら種々の事例において、本当にモルモットの体(ソーマ)がその生殖質に影響を及ぼしたのかどうかは確認されていません。既にヴァイスマンは、次のように異論を唱えています。ブラウン・セカールが手術を行う際、モルモットの体内に何らかの微生物が侵入したのかも知れない。その微生物が神経組織に栄養を摂取するための恰好の場を見付け、生殖要素にまで侵入して病気を媒介したのではないか、と。この異論はブラウン・セカール本人によって斥けられましたが、ヴァイスマンよりもっと現実的な反論をすることもできるでしょう。例えばヴォワザンとペロンは、モルモットの癲癇の発作後排泄される有毒物質を動物に注射すると、痙攣症状が惹き起こされることを実験によって明らかにしました。ブラウン・セカールがモルモットの神経に加えた損傷には栄養障害が伴いましたが、この栄養障害は、痙攣を惹き起こす毒物が形成されたことを象徴するものに違いありません。そしてこの毒物がモルモットからその精子もしくは卵にまで達し、胚が発達するにつれて胚全体に及ぶ障害を惹起するものの、成長した生体では或る特定の点にしか障害の影響が現れない、ということなのでしょう。結局これらの事例では、すべてがシャランやドラマール、ムーシュなどの実験と同じ経過を辿っているように思えます。彼らの実験によれば、妊娠中のモルモットの肝臓や腎臓を傷つけると、その損傷が子孫に遺伝します。この遺伝は、母体の器官が傷つけられることで生じた特殊な「細胞毒素」が、単に損傷した母体の器官に対応する胎児の器官に作用することによって惹き起こされたものに過ぎません。同じ生理学者達が以前行った観察においてもそうであったように、これらの実験においても、確かに毒素は既に胚の段階を脱した胎児にしか影響を及ぼしません。しかしシャランが行った別の研究では、同じやり方で精子や卵にも同じ結果が生じ得ることが明らかになっています。このように見てくると、ブラウン・セカールが行った実験における獲得形質の遺伝も、胚の中毒によって説明し得ることがわかります。彼が行った実験において、障害の範囲はなるほど体の特定の場所に限定されているように見えるものの、それは例えば、アルコールによる障害と同じ過程で遺伝するものと推測されます。ところで獲得された特性が遺伝する過程は、すべてこれと同じなのではないでしょうか。
事実、獲得形質が遺伝すると考える人もそうでない人も、次のことを認めざるを得ない点では一致しています。それは、例えばアルコールの影響のような或る種の影響は、生物のみならずその生殖質にも同時に作用を及ぼすということです。そうした影響を受けると、障害が遺伝します。と言うより、実際には生殖細胞と体(ソーマ)がどちらも同じ原因の作用を受けただけなのに、あたかも親の体(ソーマ)が自己の生殖細胞に作用した(結果、障害が遺伝する)かのように見えるのです。しかしそれは一旦措き、獲得形質が遺伝すると考える人々の主張するように、体(ソーマ)は生殖細胞に影響を及ぼし得ると仮定してみましょう。その場合にも、事態の推移は、生殖細胞と体(ソーマ)がともに同じ原因の作用を受ける、と想定した場合と何も変わらないのではないでしょうか。つまり体(ソーマ)はどう頑張っても、生殖細胞の全面的な変質以上のものを惹き起こすことはできない、と考えるのが最も自然ではないでしょうか。もしそうなら、親と同じ変化が子に現れるのは例外的なことであり、言わば偶然的なことだということになります。アルコールによる障害の遺伝に関しても、事情は同じです。アルコールによる障害は確かに父から子に遺伝しますが、伝わる形は子によってまちまちであり、いずれの子の障害も父の障害に似ていないことさえあります。生殖質に現れる変化を、仮にCと名付けましょう。Cは肯定的(ポジティブ)なものでもあり得ますし、否定的(ネガティブ)なものでもあり得ます。言い換えると、何らかの物質の獲得を表している場合もあれば、その喪失を表している場合もあります。しかしどちらにしろ、結果がその原因を正確に再現することはありません。体(ソーマ)の或る部分に生じた変化によって惹き起こされた生殖細胞の変化が、形成中の新しい有機体の同じ部分に同じ変化を惹き起こすのは、その有機体の他のすべての部分がCに対して一種の免疫を持っている場合に限られます。つまり新しい有機体において同じ部分が変化するのは、形成中のその部分だけが新たに影響を受けるからでしかありません。――しかし(偶然)そうして同じ部分が変化したとしても、その部分は、それに対応する親の有機体の部分とは全く別の方向に変化することもあり得ます。
そこでわたしたちは、偏差の遺伝と形質の遺伝との区別を導入するよう提案したいと思います。個体が新しい形質を獲得すると、それによって個体は自分自身の形態から逸脱し、またその個体が保持している胚、より厳密に表現すれば減数分裂した生殖細胞(胚細胞)が発達するにつれ再生産する筈であった形態から逸脱します。この変化(逸脱)が生殖細胞を変化させる物質を生み出さない限り、或いは生殖細胞からその諸要素の幾分かを奪い取るような栄養にかかわる機能の全面的な障害を惹き起こさない限り、それは子孫に何の効果も及ぼしません。ほとんどの事例は、これに該当します。逆にこの変化が何らかの効果を及ぼすことがあるとすれば、それは恐らく、その変化が生殖質のうちに惹き起こした化学的変化を介して作用する場合です。そうした化学的変化によって、生殖細胞から発達してくる有機体のうちに元の変化が再現されることもあるかも知れません。しかしそれは例外で、多くの場合、元の変化とは別の変化が現れます。つまり、子の有機体は親の有機体と同程度に正常な形態から逸脱するものの、その逸脱の仕方はそれぞれ異なっている、ということです。子の有機体は逸脱を受け継ぐのであって、形質を受け継ぐのではありません。したがって一般に、或る個体の獲得した習慣は、恐らくその子孫に何の影響も与えません。仮に影響を与えることがあるにしても、子孫に現れる変化と元の変化との間に目に見えるほどの類似があるとは限らない、と推測することができます。少なくともわたしたちにとって最も真実らしいと思えるのは、以上のような仮説です。いずれにせよ、この仮説に対する反証が出てこない限り、もしくは或る著名な生物学者(ジアール)が要求しているような決定的な実験が行われない限り、わたしたちは現在の観察の成果で満足する他はありません。最後に念のため獲得形質の遺伝説に最大限譲歩して、いわゆる獲得形質は、大抵の場合、生得的な形質が多少遅れて発達したものに過ぎないのではないか、という疑問を一旦念頭から消し去ってみましょう。それでもなお、諸々の事実は、獲得形質の遺伝は例外的なものであって、原則的なものではないということをわたしたちに示します。獲得形質が遺伝することで、眼のような器官が発達するなどということをどうして信じることができるでしょうか。繊毛虫類の眼点から、軟体動物の眼や脊椎動物の眼に移行するためには、厖大な数の変異がすべて同じ方向に次々に蓄積されていかなければなりません。そう考えると、個体の努力が一つ一つの偏差を生み出すことが仮にできたとしても、獲得された特性が遺伝することでどうして偏差が一定の方向に蓄積されていくのか、という疑問がやはり残ります。わたしたちとしては、新ラマルク主義も他の進化学説と同様、問題の解決をもたらすものではない、と結論せざるを得ません。
●議論の結果
さて、現代の進化思想のうち、主な進化学説をひと通り検討してきましたが、どの学説をとっても同じ乗り越え難い困難にぶつかることがわかりました。だからと言ってわたしたちは、それらすべてを否定し去るつもりは全くありません。それらの学説はいずれも多くの事実に裏付けられており、それなりの真実を含んでいる筈です。それら一つ一つが、進化過程に対する一つの観点に対応しているに違いありません。そもそも或る理論が科学的であるためには、言い換えると、細部の研究に明確な方向を与えるためには、飽くまで特定の観点に執着し続ければなりません。とは言えそれぞれの学説が手にしているのは実在の部分的な眺めでしかなく、実在はそれらすべてを超越しています。この実在こそ、哲学に固有の対象です。そして哲学は少しも応用を目指すものではない以上、科学的な精密さに囚われる必要もありません。そこで、現代の三つの主要な進化学説がそれぞれこの問題の解決に貢献していると思われるポジティブな面は何か、それらが見落としているものは何か、ということを簡単に振り返った上で、わたしたち自身の見解として、進化過程についてのより包括的な観念、より多くを含むがゆえに流動性に富んだ観念を得るためには、これら三つの努力をどこに収斂させるべきかを示したいと思います。
新ダーウィン主義者によれば、変異の本質的な原因は個体が保持している胚に内在する諸々の偏差であって、各個体の普段の行動ではありません。この点に関する限り、恐らく彼らの言い分は正しいでしょう。わたしたちが同意し難いのは、彼らが胚に内在する偏差を純粋に偶然的、かつ個体的なものと看做している点です。わたしたちの考えでは、それらの偏差は個体を介して胚から胚へと受け継がれる一つの推力が発達したものです。したがって偏差は単なる偶然的なものではなく、同一の種のすべての成員において、或いは少なくとも一部の成員において、同じ形で同時に現れるものである筈です。そしてこの点については、既に突然変異説がダーウィン主義に重要な修正を加えています。突然変異説によれば、一定以上の期間を経た種では、或るとき突然、種全体が変化しようとする傾向が顕著になるのだと言います。したがって突然変異説においては、変化しようとする傾向は偶然的なものではありません。ただしド・フリースが主張するように、突然変異が種の様々な成員において、様々な方向に起きるものだとすれば、(新ダーウィン主義と同様、突然変異説においても)変異そのものは偶発的なものだということになります。しかしそう断定するためには、まずこの説がオオマツヨイグサ以外の多くの植物種にも当て嵌まるものかどうかを確認しなければなりません(ド・フリースはオオマツヨイグサについてしかこの説を検証していません)。次に、この後の章でも説明しますが、動物の変異に比べ、植物の変異の方が偶然の占める割合が大きい点を考慮する必要があります。何故なら植物界では、動物界ほど機能が形態に密接に依存していないからです。いずれにせよ新ダーウィン主義も、突然変異の周期が(偶然的なものではなく)或る程度決定し得るものである、という論調に傾きつつあります。したがって少なくとも動物においては、突然変異の方向も決定可能である、ということになるでしょう。問題は、その決定性がどの程度のものかということです。
わたしたちはこうして、アイマーの提唱したような仮説に辿り着きます。アイマーの考えによれば、様々な形質の変異は世代から世代へ一定の方向に継続されます。この仮説は、アイマー自身が想定している範囲内であれば認めてもよいかも知れません。言うまでもないことですが、有機的世界の進化全体があらかじめ決定されている、などということはありません。寧ろ逆に、有機的世界においては形態が次々に創造されていくところに生命の自発性が現れています。しかしこの非決定性は、完全なものではあり得ません。この非決定性のどこかに決定性の入り込む余地がある筈です。例えば眼のような器官は、紛れもなく、一定の方向への連続的な変異によって形成されたものでしょう。そうでなければ、歴史を全く共有していない幾つかの種において、構造の類似した眼が出現することをどうやって説明したらよいのか見当がつかなくなります。しかしこの(眼の類似という)結果は、物理・化学的な諸々の原因の組み合わせによって確実に生じ得る、とアイマーが主張するとき、わたしたちは彼と袂を分かちます。様々な仮説の検証に好適な眼の例についてわたしたちが立証しようとしたのは、それと反対のこと、すなわち、もし「定向進化」があるとすれば、そこには心理的な原因が介入してくる筈だ、ということです。
一部の新ラマルク主義者が依拠しているのが、まさにこの心理的な性質の原因です。わたしたちの考えでは、新ラマルク主義の最も評価すべき点はここにあります。しかしこの原因が個体の意識的努力に過ぎないならば、それが作用し得る範囲はかなり限られます。そういった原因が介入し得るのは動物界だけで、植物界においてそれが作用することはないでしょう。さらに動物においても、それが作用するのは直接または間接に意志が影響を及ぼし得る幾つかの点に限られるでしょうし、またよしんば作用するにしても、器官の複雑さを増大させるような深い変化を惹き起こすことができるとは到底思えません。そういうことが可能なのは、獲得形質が規則的に遺伝し、蓄積されていく場合だけです。ところが獲得形質の遺伝は原則的なものではなく、例外的なものであるというのが真実であるように思われます。遺伝的変化が一定の方向に累積され、組織されながら次第に複雑になっていく器官を形成する場合、そうした変化が或る種の努力と結び付いているのは紛れもない事実でしょう。ただしそれは、個体の努力よりも遥かに深く、環境からも独立している努力、同じ種の成員のほとんどが共有している努力、それら成員の体(ソーマ)に内属するだけでなく、個体の保持する胚に内属する努力、まさにそのために子孫に確実に伝わる努力です。
●エラン・ヴィタール
こうしてわたしたちは大きく回り道をした末に、わたしたちの出発点となったエランという観念、すなわち胚と胚とを結ぶ連結符とも言うべき成体となった有機体を介して、世代から世代へと伝えられていく生命の根源的な弾みという観念に連れ戻されます。諸々の進化系統に分散しながらも存続するこの弾みは、変異の根本的な原因、少なくとも規則的に遺伝し、累積され、新しい種を創造する変異の根本的な原因です。一般に共通の幹から分岐した種は、進化すればするほどより一層分岐する傾向を強めていきます。しかし共通の弾みという仮説が正しいなら、それらの種は幾つかの点では同じように進化し得る筈であり、同じように進化しなければなりません。残された課題は、わたしたちが選んだ例、軟体動物と脊椎動物の眼の形成の例でこのことをより明確に示すことです。それによって「根源的な弾み」とはどういうものか、より一層明らかにすることができるでしょう。
眼のような器官を観察すると、対照的な二つの点、すなわち構造の複雑さと機能の単純さに同時に驚かされます。眼は鞏膜、角膜、網膜、水晶体などそれぞれ異なる部分で構成されていますが、それら各部分もまた、無数の部分に細分することができます。網膜だけを取ってみても、周知のようにそれは神経節細胞、双極細胞、視細胞という三つの神経細胞の層が積み重なったものであり、それぞれ独自の個性を持った各層が極めて複雑な有機体を形成しています。しかし今辛うじて示し得たのは、網膜の精緻な構造の簡略的な図式に過ぎません。眼を機械に喩えると、眼という機械は、それ自体が極めて複雑な無数の機械で構成されている、と言うことができます。一方、見るという行為は一つの単純な事実です。眼を開けば、それだけで視覚が働きます。機能が単純であるだけに、無限に複雑なこの機械を組み立てるに当たって、自然がほんの少しでも手を抜けば視覚は不可能になってしまうに違いありません。器官の複雑さと機能の単純さとのこの際立った対照は、わたしたちの精神を当惑させます。
機械論は、この眼という機械が外的環境の影響の下に次第に形成されてきた、とわたしたちに教えます。外的環境は組織に直接的に作用する場合と、適者を選択することによって間接的に組織に作用する場合があります。しかし機械論がどんな形を取るにせよ、また機械論は、或る程度までは細部の説明に役立つのは事実であるにせよ、眼という機械の部分相互の相関関係についてはわたしたちに何も教えてくれません。
そこで、目的論の出番となります。目的論によれば、機械の諸部分は、あらかじめ立てられた計画に基づき、何らかの目的のために寄せ集められたものだと言います。この点で、目的論は自然の働きを職人の仕事と同一視しています。職人は確かに、或る観念を実現するために、また或るモデルを模倣するために、諸部分を集めることによって事を進めます。それゆえ機械論が、目的論の持つ擬人論的な性格を非難するのは間違いではありません。しかし機械論は、自分自身同じ方法に従っていること、目的論の辿った道筋を、一部を除きそのままなぞっているに過ぎない、ということを見落としています。追求すべき目的とか、手本となるモデルといった観念を機械論が排除したのは事実ですが、自然は職人と同様、諸部分を寄せ集めることによって事を進める、と考える点では機械論も目的論と変わりがありません。胚の成長過程を少しでも観察すれば、生命の作用は、そういう(諸要素の寄せ集めという)やり方とは全く異なっていることにすぐさま気付く筈です。生命は諸要素の結合や累積によって進展するのではなく、分離と分裂によって進展します。
したがって機械論の観点と目的論の観点は、どちらも乗り越えられなければなりません。実を言うと、両者はいずれも、人間の精神(知性)が自分の仕事を省みることによって自ずと導かれた観点に過ぎません。しかしそれらを乗り越えると言っても、どの方向を目指せばよいのでしょうか。先に述べたように、或る器官の構造を分析していくとどこまでも細かく分解できるのに引き換え、その機能は極めて単純です。器官の無限の複雑さと機能の極端な単純さのこの対照こそ、問題解決への糸口となるに違いありません。
一般に、或る対象が一面では単純に見え、他面では無限に多くの要素によって構成されているように見えるとき、これら二つの面は、同じ重要性、と言うより同程度の実在性を持つわけではありません。単純さが対象そのものに属しているとすれば、無限の複雑さは、わたしたちが対象の周囲を回りながらその対象を捉える観点に、別の言い方をすると、わたしたちの感覚や知性がその対象を表象する際に並列する記号に、より一般化して言えば、わたしたちが対象を模倣しようとする際に用いる諸要素に、ただし対象とは秩序の異なる諸要素に属しています。対象はそれらの諸要素とは本性が異なる以上、両者の間に共通の尺度は存在し得ません。絵画の巨匠がカンヴァスに描いた絵を思い浮かべてみましょう。わたしたちは様々な色彩のモザイクの小片を用いて、その絵を模倣することができます。モザイクの小片が小さければ小さいほど、また小片の数が多ければ多いほど、そして小片の色調が多彩であればあるほど、原画の曲線やニュアンスをより正確に再現することができるでしょう。しかしその巨匠が頭の中で単純なものとして思い描き、渾然一体のものとしてカンヴァスに定着させようとしたイメージ、不可分な直観の投影として現れれば現れるほど完璧なものに近付いていくイメージと完全に等しいものを得ようと思えば、無限に小さく、無限のニュアンスを持った無数の要素が必要になります。さてここで、わたしたちの眼は、この巨匠の作品がモザイクの効果としか見えないように作られていると想定してみましょう。或いはわたしたちの知性は、カンヴァスに描かれた絵をモザイクによる創作としてしか理解できないように出来ていると想定してみましょう。その場合、わたしたちは寄せ集められたモザイクの小片の堆積以外のものについて語ることができず、機械論的な仮説に依拠することになります。或いは作品の創作には、寄せ集められた物質的な材料に加えて、モザイク師が仕事を進めるための計画が必要であった、と主張することもできるでしょう。その場合には、わたしたちは目的論の立場を取ることになります。しかしどちらの立場を取るにせよ、わたしたちが創作の真の過程に辿り着くことはありません。何故なら、そもそも寄せ集められた小片などどこにも存在していないからです。存在するのは絵そのもの、つまりカンヴァスに投影された単純な行為であって、わたしたちがそれを知覚したというただそれだけの理由によって、その行為はわたしたちの眼に対して幾千もの小片に分解されます。分解された小片の配列が驚くべきものに見えるのは、小片が再構成されることでその配列が出来上がった、という風に考えてしまうからです。同様に、複雑極まりない構造を持った眼も、見るという単純な行為以外のものではありません。その単純な行為が自ら分解してわたしたちにとって細胞のモザイクとなり、モザイク全体が一つの集合体として表象される結果、その秩序が奇跡的なものに見えてくる、というわけです。
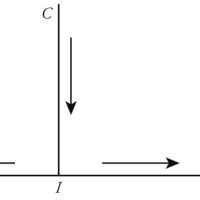
わたしが点Aから点Bまで手を挙げるとき、この運動はわたしにとって同時に二つの形を取って現れます。内からこの運動を感じる場合、それは単純で不可分な行為として現れます。外からこの運動を見る場合、それは一つの曲線ABとして現れます。その曲線上に、わたしは好きなだけ多くの位置を区別することができ、またその曲線そのものも、それらの位置が何らかの秩序に従って相互に配列されたものとして定義することができます。しかし無数の位置にせよ、或いはそれらの位置を相互に結び付ける秩序にせよ、点Aから点Bに手を動かす不可分な行為から自動的に生じたものに過ぎません。機械論がそれらの位置に着目するのに対し、目的論は位置の秩序を考慮に入れます。その反面、機械論と目的論は、実在そのものである運動を揃って見逃してしまいます。運動は、或る意味では諸々の位置やその秩序以上のものです。何故なら運動が与えられさえすれば、それが不可分で単純なものである限り、継起する無数の位置やその秩序も同時に与えられ、それに加えて位置でも秩序でもない本質的な或るもの、すなわち動性が与えられるからです。しかし別の意味では、運動は一連の位置やそれらを結び付ける秩序以下のものです。何故なら諸々の点を或る秩序に従って配列するためには、まずその秩序を表象し、次いで諸々の点でもってこの秩序を実現しなければならないからです。つまり諸々の要素を寄せ集める作業と、知性が必要だからです。ところが現実には、手の単純な運動にはそういったものは全く含まれていません。手の運動は、人間的な意味では知性的なものではありませんし、諸要素から構成されているわけではない以上、それらの寄せ集めでもありません。眼と視覚との関係についても同じことが言えます。視覚には、眼を構成する諸々の細胞や、それら相互の結び付き以上のものがあります。この意味で、機械論も目的論も不十分です。が、別の意味では機械論と目的論は行き過ぎです。何故なら機械論も目的論も、無限に複雑な無数の要素を視覚という単純な行為に仕上げるという、ヘラクレスに命じられたどんな難題よりも難しい仕事を自然に割り当てていますが、自然は眼を作るのに、(ヘラクレスのような怪力どころか)わたしが手を挙げるのと同じ程度の労力しか必要としなかったからです。自然の単純な行為は自動的に無数の要素に分解されるのであって、それらの要素が同じ観念の下に秩序づけられていることにわたしたちは事後的に気付くに過ぎないのです。ちょうどわたしの手の運動が無数の点を(その軌道上に)残していった後で、初めてそれらの点が同じ一つの方程式を満たしていることに気付くように。
しかし以上のことを理解するのは、決して易しいことではありません。というのもわたしたちは、有機的組織化を一種の製作として表象せずにはいられないからです。しかし(そういったわたしたちの先入観に反して)、製作と有機的組織化とは飽くまで別のものです。まず製作に関して言えば、それは人間に固有の活動です。製作とは、材料を様々な部分に分割した上で、部分同士を互いに組み合わせて一つの共通の機能が得られるようにそれらを寄せ集めることです。寄せ集めた諸部分を、わたしたちはその機能の周囲に、すなわち、それら諸部分の言わば中心を占めると想定された機能の周囲に並べていきます。したがって製作は、周辺から中心に、哲学者の表現に従えば多から一に進みます。他方、有機的組織化の働きは中心から周辺へと進みます。それはほとんど数学的な点と言ってよい一つの点から始まり、その点を中心として、徐々に大きさを増していく同心円状の波動によって周辺に拡がっていきます。また、扱う材料が多ければ多いほど効果が増していく製作の働きは、集中と圧縮によって進められます。反対に有機的組織化の作用には爆発を髣髴とさせるものがあります。それは出発点においては、極めて小さい場所と最小限の材料しか必要としません。有機的組織化の力は(本来自己の外に出ることを潔しとせず)、不承不承空間の中に入っていった、という印象を受けます。事実、胚生の過程において原動力となる精子は、有機体の中で最も小さい細胞の一つです。しかも胚生の過程と直接かかわりがあるのは、精子の極く一部でしかありません。
とは言え上述した製作と有機的組織化との相違は、表面的な差異に過ぎません。それらの差異を掘り下げていけば、もっと深い差異が見つかる筈です。
製作されたもの、すなわち製作物は、製作という作業の形式を描いています。ということは、製作者はその製作物のうちに、彼が注ぎ込んだだけのものを再発見することができる、ということです。例えば一つの機械を作ろうとするとき、製作者は機械の部品を素材から一つずつ切り出し、次いでそれらを寄せ集めます。完成した機械のうちには、部品の一つ一つと、部品の総体を見て取ることができるでしょう。ここでは結果の総体が作業の総体を表しており、作業の各部分には結果の或る一部分が対応しています。
実証科学は、有機的組織化を製作と同じ種類の仕事と看做すことができ、またそう看做さなければならないことはわたしも認めます。そうした条件においてのみ、実証科学は有機体の研究に着手することができます。実際、実証科学の目的は事物の基礎を明示することではなく、事物に働きかける最良の手段を提供することです。ところで、物理学と化学は既に学問として確立されています。そしてわたしたちが生ある物質に働きかけることができるのは、それが物理学や化学の方法で扱える場合に限られます。したがって有機的組織化を科学的に研究するためには、有機体をまず機械に見立てなければなりません。そこで細胞は機械の部品と看做され、有機体はそれらの部品の集合体と看做されることになるでしょう。さらに各部分を有機的組織化する部分的な働きは、全体を有機的組織化する働きの実在的な要素と看做されます。こうした観点が科学の観点です。わたしたちの考えでは、哲学の観点はそういったものとは全く異なります。
なるほど、有機的組織化された機械(器官)の全体は有機的組織化の働きの全体を表していると言ってよいかも知れません(と言っても近似的にそう言えるに過ぎません)が、この機械(器官)の各部分は、(有機的組織化という)作業の各部分には対応していません。というのもこの機械(器官)の物質性が表しているのは、製作物の場合とは違い、用いられた手段の全体ではなく、回避された障害の全体だからです。それは肯定的な実在と言うよりも、寧ろ一つの否定です。前著で示したように、理論上、視覚はわたしたちの目の届かない無数の事物にも達し得る能力です。ところがそのような視覚は、行動にまで延長されることは(なく現実のものになることも)ありません。したがってそれ(理論上の視覚)は、生物よりも寧ろ亡霊に相応しい視覚です。生物の視覚は有効なものでなければならず、生物が働きかけ得る対象に範囲が限定されています。生物の視覚とは運河の開鑿のごときものであり、視覚器官は運河を開鑿する作業を象徴しているに過ぎません。運河の開鑿は、両岸に運び出され、積み上げられた土によって説明することはできないように、視覚器官の創造は解剖学的要素の集積によって説明することはできません。機械論者ならば、運河は土を積んだ荷車を一台ずつ運び出して造られたのだ、と説明し、目的論者はそれに付け加えて、土は計画もなく運び出されたのではなく、労働者が一つの計画に従って運び出したのだ、と説明するでしょう。しかし機械論者の説明も目的論者の説明も、どちらも見当違いと言う他はありません。何故なら運河は土(障害物)の運搬作業や積み上げ作業とは別のやり方で造られたものだからです。
わたしたちは先ほど、自然が眼を形成する過程を手を挙げるという単純な動作になぞらえましたが、運河の比喩よりも、手の比喩の方が的確です。ただその際わたしたちは、手は何の抵抗にも出会わないと想定しました。そこで今度は、わたしの手は空気中を動くのではなく、鉄屑の中を進んでいくものとしましょう。わたしが手を前へ進めれば進めるほど鉄屑は圧縮されて抵抗を増していき、或る瞬間が来ると、わたしの手は最早それ以上先へは進めなくなります。ちょうどその瞬間、鉄屑は或る一定の形、すなわち停止した手と腕の一部の形そのままに配列され、秩序づけられます。このとき手と腕が仮に透明であったとすると、それを見ている人は、鉄屑がそのように配列されている理由を(手の動きに求めるのではなく)、鉄屑そのものうちに、或いは鉄屑の堆積に内在する力に求めるに違いありません。或る人は鉄屑一つ一つの位置を、隣接する鉄屑がそれに及ぼす作用と関連付けるでしょう。そういう人は機械論者です。また別の人は、全体的な一つの計画が鉄屑の個々の作用を細部に至るまで支配しているに違いない、と考えるでしょう。そういう人は目的論者です。しかし実際には、そこには単に一つの不可分な行為、手が鉄屑の中を進んでいくという行為があったに過ぎず、鉄屑の筆舌に尽くし難い動きの一つ一つやその最終的な配列の秩序は、この不可分な手の運動を言わば否定的に表現しているに過ぎません。それは抵抗の全体的な形であって、鉄屑の個々の積極的作用の総合ではありません。鉄屑の配列を「結果」と呼び、手の運動を「原因」と呼ぶとすれば、結果の全体は原因の全体によって説明される、とは言い得るでしょうが、原因の各部分に結果の各部分が対応することは決してないでしょう。この場合、機械論の説明も目的論の説明も用をなさず、独自の説明の仕方を見つける必要があります。例えばわたしたちが提示する仮説では、視覚と視覚器官との関係は、おおよそ手と鉄屑との関係に、すなわち手と、手の運動を描き、手の運動によって運河を掘られ、手の運動を限定する鉄屑との関係になぞらえることができます。
手の努力が大きければ大きいほど、手はそれだけ鉄屑の奥まで進むことができます。そして手がどこで止まるにせよ、鉄屑は瞬間的かつ自動的に均整が取れ、相互に調整されます。視覚と視覚器官との関係においても事情は同じです。視覚を構成する不可分な行為がどこまで進むかに応じて、視覚器官の物質性をなす要素、相互に調整された要素が多くなったり少なくなったりします。にもかかわらずその秩序は、どんな場合にも完全無欠です。それが不完全であることは決してありません。何故ならもう一度言いますが、秩序を生み出す実在的な過程には部分が存在しないからです。これこそ、機械論者と目的論者がともに見逃している点であり、わたしたちが眼のような器官の精妙な構造に驚く際に見逃している点です。わたしたちの驚きの奥底には、実現されたのは全体の秩序の一部だけであろう、という思い込み、完全な秩序の実現は一種の恩寵である、という思い込みが常に潜んでいます。目的論者は、この恩寵が目的因によって一度に与えられる、と考えます。機械論者は、この恩寵が自然淘汰によって少しずつ与えられる、と考えます。しかしどちらも秩序のうちに肯定的なものを見ていることに変わりはなく、したがって両者はいずれも、秩序を生み出すその原因のうちに分割可能なもの、あらゆる可能な完成度を許容するものを見ています。実際には、(生命の領域では)原因の強さに程度の違いがあっても、その結果はひと纏まりのものとして、完全な形でしか与えられません。この原因が視覚の方向にどの程度進んでいるかに応じて、下等有機体の単なる色素群が生み出されたり、セルプラ(環形動物)の原基的な眼が生み出されたり、アルキオパ(同じく環形動物)の組織の分化が見て取れる眼が生み出されたり、鳥の驚くほど完成された眼が生み出されたりします。これらの器官は複雑さの程度は違えど、常に同じように調整されています。そういうわけで、二つの異なった動物種の系統がどんなにかけ離れていようが、双方の視覚への歩みが同じ程度に進んでいる限り、どちらの種にも同じような視覚器官が見られるでしょう。何故なら器官の形態は、機能がどの程度発達したかを示す尺度でしかないからです。
しかし視覚への歩みなどという言葉を持ち出すと、目的性という古い考え方に逆戻りすることにはならないでしょうか。もしこの歩みが、意識的なものか無意識的なものかを問わず到達すべき目標を表すものならば、確かにその通りかも知れません。しかし実際には、この歩みは生命の根源的なエランによって行われ、その運動そのものの中に含まれています。だからこそ、この歩みは互いに独立した進化系統の上に見出されるのです。ところでこの歩みは何故、またどのようしてこの運動の中に含まれるのでしょうか。そう問われれば、わたしたちは次のように答えるでしょう。生命とは、何よりもまず生(なま)の物質に働きかける(つまり行動しようとする)一つの傾向である、(だからこそそれはその運動の中に含まれ、そのような傾向としてその運動の中に含まれるのだ)と。この働きの方向は、恐らく前もって決定されているわけではありません。そこから、生命が進化の途上撒き散らしていった形態の予測不可能な多様性が帰結します。この働きは、程度の違いこそあれ常に偶然性を帯びています。或いは偶然性とは言わないまでも、少なくとも選択の兆しを含んでいます。ところで選択は、幾つかの可能な行動をあらかじめ表象することを前提としています。したがって行動そのものに先立って、行動の様々な可能性が生物に素描されていなければなりません。視覚的知覚とはそれ以外のものではありません。目に見える物体の輪郭は、それらに対するわたしたちの可能的行動の素描です。視覚が様々な程度で、極めて多くの動物に見られるのはそのためです。視覚は、それが同じ強度に達するところではどこでも、同じ複雑さを持つ構造を通して姿を現すでしょう。
わたしたちは一般に器官の構造の類似について、その中でも特に眼の構造の類似について論じてきました。というのも一方で機械論に対して、他方で目的論に対して、わたしたちの態度を明確にしなければならなかったからです。次に、わたしたちの態度そのものをより正確に記述しなければなりません。次章において、わたしたちは進化の多岐にわたる成果、ただし今度はそれらの類似している点ではなく、それらが相互に補完し合っている点を検討することによって、わたしたちの考えをより明確にしていきたいと思います。
(つづく)
周知のようにラマルクは、生物にはその器官を使用するかしないかに応じて変異し、しかもそうして獲得した変異を子孫に伝える能力が備わっている、と主張しました。今日においても、ラマルクの思想の流れを汲む学説は一定数の生物学者に支持されています。この学説によれば、新しい種を生み出す変異は、胚そのものに内在する偶発的なものでもなければ、一定の形質を、有用性とは無関係に一定の方向に発達させる一種独特な決定論に従うものでもありません。それは、生物が自分に課せられた生存条件に適応しようとする努力そのものから生まれます。無論この努力は、外的環境の圧力によって機械的に惹起される或る器官の機械的作用に過ぎない場合もあるでしょうが、意識と意志を含む場合もあるでしょう。この学説を代表する人物の一人であるアメリカの博物学者コープは、努力を後者の意味に解しています。したがって新ラマルク主義そのものは必ずしも内的で心理的な発達原理に訴える必然性は持たないとしても、この学説は現在の進化思想のうち、そうした発達原理を受け入れ得る唯一の学説と言えます。また新ラマルク主義は、それぞれ独立した複数の発達系統において、同一の複雑な器官が形成されることを説明できそうな唯一の学説でもあります。事実、同じ環境を利用しようとする同じ努力が、同じ結果に達するのは自然なことです。さらに外的環境の提出する問題が唯一つの解答しか許さないような性質のものであれば、同じ努力が同じ結果に達する蓋然性はより一層高くなる、と言えるでしょう。とは言え検討すべき問題が一つ残されています。それは、この「努力」という言葉には、どんな新ラマルク主義者が考えているよりももっと深い心理的な意味を賦与しなければならないのではないか、ということです。
実際、単なる大きさの変異と、形態の変化とは区別して考えなければなりません。或る器官の機能を働かせ続ければ、それが強化され大きさを増すことを否定する人はいないでしょう。しかしそのことと、例えば軟体動物の眼や脊椎動物の眼が漸進的に発達することとは全く別のことです。眼の発達の原因を受動的に受け取られた光の影響に求めるならば、たった今批判したばかりの説に逆戻りすることになります。逆に内的作用を持ち出すのであれば、わたしたちが通常努力と呼んでいるものとは全く別のものを問題にする必要があります。何故なら繊毛虫類の眼点から脊椎動物の眼に移行するまでの間には、驚くほど整然とした無数の複雑さの段階が存在するにもかかわらず、努力によって或る器官がほんのわずかでも複雑になったなどという話は聞いたことがないからです。動物に関する限り、進化過程を努力によるものと看做すそうした考え方を認めてもよい知れません。しかしこの考え方を、どうして植物界にまで拡げることができるでしょうか。植物界では、形態の変異は常に機能の変化を含んでいるようには見えませんし、それを伴っているようにも思えません。変異の原因を心理的な性質のものと仮定するのはよいとしても、その原因を努力と呼ぶためには、言葉の意味を異常に拡張しなければならないでしょう。わたしたちがすべきことはこの努力そのものを掘り下げ、もっと深い原因を探ることなのです。
特に規則的に遺伝する変異が問題となる場合には、そういった探究が一段と重要になってきます。わたしたちは、ここで獲得形質の遺伝に関する論争の詳細に立ち入るつもりはありません。わたしたちの専門外の問題について、余り明確な態度を表明するのは本来なら差し控えたいところです。とは言っても、この問題に全く無関心でいるわけにもいきません。今日、哲学者は漠然とした一般論で満足しているわけにはいかないこと、哲学者も科学者の行った実験の細部を辿り、その結果について科学者と意見を交換する義務があること、このことを最も痛感するのが、他でもない、獲得形質の遺伝に関する問題を扱うときです。仮にスペンサーが獲得形質の遺伝に関する問題を最初に自らに課していたとすれば、恐らく彼の進化論は全く違ったものになっていたに違いありません。個体が身に付けた習慣が、極めて例外的な場合にしか子孫に遺伝しない(わたしたちにはそう思えるのですが)ならば、スペンサーの心理学は全面的に修正されなければならないでしょうし、彼の築いた哲学の大部分はその根拠を失ってしまう筈です。そういうわけで、獲得形質の遺伝に関する問題がどのように提起されているか、またこの問題の解決をどの方向に求めるべきかについて、わたしたちの見解を率直に述べて置くことにします。
獲得形質が遺伝するという説はかつて独断的に肯定されていましたが、その後、生殖細胞に備わっているとされる性質からア・プリオリに引き出された理由によって、同じく独断的に否定されるに至りました。ヴァイスマンが、生殖質は連続するという仮説に基づき、生殖細胞――卵と精子――が体細胞からほとんど独立していると結論するに至ったのは周知の通りですが、そこから獲得形質は遺伝しないという主張が広がっていったのです。この主張は、今日においても最も有力な説の一つと言って差し支えありません。――しかし獲得形質が遺伝し得るものであることが万に一つでも実験によって明らかになった場合、生殖質は現在考えられているほど周囲の体細胞から独立していないということになり、事実上、獲得形質は遺伝すると考えてもよい、ということになるでしょう。つまり獲得形質は遺伝するか否かという机上の議論には意味がなく、問題の解決はひとえに実験にかかっている、ということになります。とは言え口で言うのは簡単ですが、難しいのはここから先の部分です。いわゆる獲得形質とは、多くの場合、習慣もしくは習慣の結果を指しますが、獲得された習慣の根底に、自然的な資質なり傾向が存在していないことは滅多にありません。そのため遺伝したのは個体の体(たい。原語はsomaで、生殖細胞を除く体全体)が獲得した習慣なのか、或いは寧ろ、獲得された習慣に先立つ自然的な傾向ではないのか、という疑問が常に付いて回ります。そうした傾向は、既に個体に内属していたのかも知れず、したがってもともとその胚に、そして個体の生殖細胞に内属していたのかも知れません。それゆえモグラの視力が悪いのは、地下で生活する習慣を身に付けたからだ、と結論するのは早計です。恐らくモグラは、眼が衰退しつつあったからこそ地下での生活を余儀なくされたのでしょう。そうだとすれば、視力が失われていく傾向は生殖細胞から生殖細胞へと遺伝したのであって、モグラの体(ソーマ)そのものは何も獲得せず、何も失わなかったと考えるのが自然です。フェンシングの名手の息子が父親よりずっと早く優れた選手になったからと言って、親の習慣が子に遺伝したと決め付けることはできません。というのもその場合、発達途上にある何らかの自然的な傾向が、父親を生み出した生殖質から息子を生み出した生殖質へと遺伝する過程で、起源にあるエランの作用によって成長した結果、言ってみれば息子は父親の身に付けた習慣など見向きもせず父親を凌駕する反射神経を獲得したのかも知れないからです。長期間にわたって行われる動物の家畜化についても、同様のことが言えます。家畜化を目的として、動物の中から或る特定の種やその種の成員の一部を選ぶ際、選定の目安となるのはそれらの持つ自然的性向ですが、遺伝するのはそれらが身に付けた習慣なのか、それとも選定の目安となった或る種の自然的性向なのかを見極めるのは困難です。実を言うと、獲得された特性が遺伝する事例のうち、疑わしい事例や、様々な解釈を容れる余地のある事実をすべて除外すると、絶対に異論を挟む余地のないものとして残るのは、何人もの生理学者によって繰り返し再現され、確認されているブラウン・セカールの有名な実験くらいしかありません。彼の実験が明らかにしたところによれば、モルモットの脊髄もしくは坐骨神経を切断すると癲癇症状が現れ、子孫にもこの症状が遺伝します。坐骨神経や索状体などの損傷は、モルモットに様々な障害を惹き起こしましたが、これらの障害は、時として最初の障害とは異なる形、例えば眼球突出や足指喪失といった形で子孫に遺伝することもあったと言います。――しかしブラウン・セカールが提示したこれら種々の事例において、本当にモルモットの体(ソーマ)がその生殖質に影響を及ぼしたのかどうかは確認されていません。既にヴァイスマンは、次のように異論を唱えています。ブラウン・セカールが手術を行う際、モルモットの体内に何らかの微生物が侵入したのかも知れない。その微生物が神経組織に栄養を摂取するための恰好の場を見付け、生殖要素にまで侵入して病気を媒介したのではないか、と。この異論はブラウン・セカール本人によって斥けられましたが、ヴァイスマンよりもっと現実的な反論をすることもできるでしょう。例えばヴォワザンとペロンは、モルモットの癲癇の発作後排泄される有毒物質を動物に注射すると、痙攣症状が惹き起こされることを実験によって明らかにしました。ブラウン・セカールがモルモットの神経に加えた損傷には栄養障害が伴いましたが、この栄養障害は、痙攣を惹き起こす毒物が形成されたことを象徴するものに違いありません。そしてこの毒物がモルモットからその精子もしくは卵にまで達し、胚が発達するにつれて胚全体に及ぶ障害を惹起するものの、成長した生体では或る特定の点にしか障害の影響が現れない、ということなのでしょう。結局これらの事例では、すべてがシャランやドラマール、ムーシュなどの実験と同じ経過を辿っているように思えます。彼らの実験によれば、妊娠中のモルモットの肝臓や腎臓を傷つけると、その損傷が子孫に遺伝します。この遺伝は、母体の器官が傷つけられることで生じた特殊な「細胞毒素」が、単に損傷した母体の器官に対応する胎児の器官に作用することによって惹き起こされたものに過ぎません。同じ生理学者達が以前行った観察においてもそうであったように、これらの実験においても、確かに毒素は既に胚の段階を脱した胎児にしか影響を及ぼしません。しかしシャランが行った別の研究では、同じやり方で精子や卵にも同じ結果が生じ得ることが明らかになっています。このように見てくると、ブラウン・セカールが行った実験における獲得形質の遺伝も、胚の中毒によって説明し得ることがわかります。彼が行った実験において、障害の範囲はなるほど体の特定の場所に限定されているように見えるものの、それは例えば、アルコールによる障害と同じ過程で遺伝するものと推測されます。ところで獲得された特性が遺伝する過程は、すべてこれと同じなのではないでしょうか。
事実、獲得形質が遺伝すると考える人もそうでない人も、次のことを認めざるを得ない点では一致しています。それは、例えばアルコールの影響のような或る種の影響は、生物のみならずその生殖質にも同時に作用を及ぼすということです。そうした影響を受けると、障害が遺伝します。と言うより、実際には生殖細胞と体(ソーマ)がどちらも同じ原因の作用を受けただけなのに、あたかも親の体(ソーマ)が自己の生殖細胞に作用した(結果、障害が遺伝する)かのように見えるのです。しかしそれは一旦措き、獲得形質が遺伝すると考える人々の主張するように、体(ソーマ)は生殖細胞に影響を及ぼし得ると仮定してみましょう。その場合にも、事態の推移は、生殖細胞と体(ソーマ)がともに同じ原因の作用を受ける、と想定した場合と何も変わらないのではないでしょうか。つまり体(ソーマ)はどう頑張っても、生殖細胞の全面的な変質以上のものを惹き起こすことはできない、と考えるのが最も自然ではないでしょうか。もしそうなら、親と同じ変化が子に現れるのは例外的なことであり、言わば偶然的なことだということになります。アルコールによる障害の遺伝に関しても、事情は同じです。アルコールによる障害は確かに父から子に遺伝しますが、伝わる形は子によってまちまちであり、いずれの子の障害も父の障害に似ていないことさえあります。生殖質に現れる変化を、仮にCと名付けましょう。Cは肯定的(ポジティブ)なものでもあり得ますし、否定的(ネガティブ)なものでもあり得ます。言い換えると、何らかの物質の獲得を表している場合もあれば、その喪失を表している場合もあります。しかしどちらにしろ、結果がその原因を正確に再現することはありません。体(ソーマ)の或る部分に生じた変化によって惹き起こされた生殖細胞の変化が、形成中の新しい有機体の同じ部分に同じ変化を惹き起こすのは、その有機体の他のすべての部分がCに対して一種の免疫を持っている場合に限られます。つまり新しい有機体において同じ部分が変化するのは、形成中のその部分だけが新たに影響を受けるからでしかありません。――しかし(偶然)そうして同じ部分が変化したとしても、その部分は、それに対応する親の有機体の部分とは全く別の方向に変化することもあり得ます。
そこでわたしたちは、偏差の遺伝と形質の遺伝との区別を導入するよう提案したいと思います。個体が新しい形質を獲得すると、それによって個体は自分自身の形態から逸脱し、またその個体が保持している胚、より厳密に表現すれば減数分裂した生殖細胞(胚細胞)が発達するにつれ再生産する筈であった形態から逸脱します。この変化(逸脱)が生殖細胞を変化させる物質を生み出さない限り、或いは生殖細胞からその諸要素の幾分かを奪い取るような栄養にかかわる機能の全面的な障害を惹き起こさない限り、それは子孫に何の効果も及ぼしません。ほとんどの事例は、これに該当します。逆にこの変化が何らかの効果を及ぼすことがあるとすれば、それは恐らく、その変化が生殖質のうちに惹き起こした化学的変化を介して作用する場合です。そうした化学的変化によって、生殖細胞から発達してくる有機体のうちに元の変化が再現されることもあるかも知れません。しかしそれは例外で、多くの場合、元の変化とは別の変化が現れます。つまり、子の有機体は親の有機体と同程度に正常な形態から逸脱するものの、その逸脱の仕方はそれぞれ異なっている、ということです。子の有機体は逸脱を受け継ぐのであって、形質を受け継ぐのではありません。したがって一般に、或る個体の獲得した習慣は、恐らくその子孫に何の影響も与えません。仮に影響を与えることがあるにしても、子孫に現れる変化と元の変化との間に目に見えるほどの類似があるとは限らない、と推測することができます。少なくともわたしたちにとって最も真実らしいと思えるのは、以上のような仮説です。いずれにせよ、この仮説に対する反証が出てこない限り、もしくは或る著名な生物学者(ジアール)が要求しているような決定的な実験が行われない限り、わたしたちは現在の観察の成果で満足する他はありません。最後に念のため獲得形質の遺伝説に最大限譲歩して、いわゆる獲得形質は、大抵の場合、生得的な形質が多少遅れて発達したものに過ぎないのではないか、という疑問を一旦念頭から消し去ってみましょう。それでもなお、諸々の事実は、獲得形質の遺伝は例外的なものであって、原則的なものではないということをわたしたちに示します。獲得形質が遺伝することで、眼のような器官が発達するなどということをどうして信じることができるでしょうか。繊毛虫類の眼点から、軟体動物の眼や脊椎動物の眼に移行するためには、厖大な数の変異がすべて同じ方向に次々に蓄積されていかなければなりません。そう考えると、個体の努力が一つ一つの偏差を生み出すことが仮にできたとしても、獲得された特性が遺伝することでどうして偏差が一定の方向に蓄積されていくのか、という疑問がやはり残ります。わたしたちとしては、新ラマルク主義も他の進化学説と同様、問題の解決をもたらすものではない、と結論せざるを得ません。
●議論の結果
さて、現代の進化思想のうち、主な進化学説をひと通り検討してきましたが、どの学説をとっても同じ乗り越え難い困難にぶつかることがわかりました。だからと言ってわたしたちは、それらすべてを否定し去るつもりは全くありません。それらの学説はいずれも多くの事実に裏付けられており、それなりの真実を含んでいる筈です。それら一つ一つが、進化過程に対する一つの観点に対応しているに違いありません。そもそも或る理論が科学的であるためには、言い換えると、細部の研究に明確な方向を与えるためには、飽くまで特定の観点に執着し続ければなりません。とは言えそれぞれの学説が手にしているのは実在の部分的な眺めでしかなく、実在はそれらすべてを超越しています。この実在こそ、哲学に固有の対象です。そして哲学は少しも応用を目指すものではない以上、科学的な精密さに囚われる必要もありません。そこで、現代の三つの主要な進化学説がそれぞれこの問題の解決に貢献していると思われるポジティブな面は何か、それらが見落としているものは何か、ということを簡単に振り返った上で、わたしたち自身の見解として、進化過程についてのより包括的な観念、より多くを含むがゆえに流動性に富んだ観念を得るためには、これら三つの努力をどこに収斂させるべきかを示したいと思います。
新ダーウィン主義者によれば、変異の本質的な原因は個体が保持している胚に内在する諸々の偏差であって、各個体の普段の行動ではありません。この点に関する限り、恐らく彼らの言い分は正しいでしょう。わたしたちが同意し難いのは、彼らが胚に内在する偏差を純粋に偶然的、かつ個体的なものと看做している点です。わたしたちの考えでは、それらの偏差は個体を介して胚から胚へと受け継がれる一つの推力が発達したものです。したがって偏差は単なる偶然的なものではなく、同一の種のすべての成員において、或いは少なくとも一部の成員において、同じ形で同時に現れるものである筈です。そしてこの点については、既に突然変異説がダーウィン主義に重要な修正を加えています。突然変異説によれば、一定以上の期間を経た種では、或るとき突然、種全体が変化しようとする傾向が顕著になるのだと言います。したがって突然変異説においては、変化しようとする傾向は偶然的なものではありません。ただしド・フリースが主張するように、突然変異が種の様々な成員において、様々な方向に起きるものだとすれば、(新ダーウィン主義と同様、突然変異説においても)変異そのものは偶発的なものだということになります。しかしそう断定するためには、まずこの説がオオマツヨイグサ以外の多くの植物種にも当て嵌まるものかどうかを確認しなければなりません(ド・フリースはオオマツヨイグサについてしかこの説を検証していません)。次に、この後の章でも説明しますが、動物の変異に比べ、植物の変異の方が偶然の占める割合が大きい点を考慮する必要があります。何故なら植物界では、動物界ほど機能が形態に密接に依存していないからです。いずれにせよ新ダーウィン主義も、突然変異の周期が(偶然的なものではなく)或る程度決定し得るものである、という論調に傾きつつあります。したがって少なくとも動物においては、突然変異の方向も決定可能である、ということになるでしょう。問題は、その決定性がどの程度のものかということです。
わたしたちはこうして、アイマーの提唱したような仮説に辿り着きます。アイマーの考えによれば、様々な形質の変異は世代から世代へ一定の方向に継続されます。この仮説は、アイマー自身が想定している範囲内であれば認めてもよいかも知れません。言うまでもないことですが、有機的世界の進化全体があらかじめ決定されている、などということはありません。寧ろ逆に、有機的世界においては形態が次々に創造されていくところに生命の自発性が現れています。しかしこの非決定性は、完全なものではあり得ません。この非決定性のどこかに決定性の入り込む余地がある筈です。例えば眼のような器官は、紛れもなく、一定の方向への連続的な変異によって形成されたものでしょう。そうでなければ、歴史を全く共有していない幾つかの種において、構造の類似した眼が出現することをどうやって説明したらよいのか見当がつかなくなります。しかしこの(眼の類似という)結果は、物理・化学的な諸々の原因の組み合わせによって確実に生じ得る、とアイマーが主張するとき、わたしたちは彼と袂を分かちます。様々な仮説の検証に好適な眼の例についてわたしたちが立証しようとしたのは、それと反対のこと、すなわち、もし「定向進化」があるとすれば、そこには心理的な原因が介入してくる筈だ、ということです。
一部の新ラマルク主義者が依拠しているのが、まさにこの心理的な性質の原因です。わたしたちの考えでは、新ラマルク主義の最も評価すべき点はここにあります。しかしこの原因が個体の意識的努力に過ぎないならば、それが作用し得る範囲はかなり限られます。そういった原因が介入し得るのは動物界だけで、植物界においてそれが作用することはないでしょう。さらに動物においても、それが作用するのは直接または間接に意志が影響を及ぼし得る幾つかの点に限られるでしょうし、またよしんば作用するにしても、器官の複雑さを増大させるような深い変化を惹き起こすことができるとは到底思えません。そういうことが可能なのは、獲得形質が規則的に遺伝し、蓄積されていく場合だけです。ところが獲得形質の遺伝は原則的なものではなく、例外的なものであるというのが真実であるように思われます。遺伝的変化が一定の方向に累積され、組織されながら次第に複雑になっていく器官を形成する場合、そうした変化が或る種の努力と結び付いているのは紛れもない事実でしょう。ただしそれは、個体の努力よりも遥かに深く、環境からも独立している努力、同じ種の成員のほとんどが共有している努力、それら成員の体(ソーマ)に内属するだけでなく、個体の保持する胚に内属する努力、まさにそのために子孫に確実に伝わる努力です。
●エラン・ヴィタール
こうしてわたしたちは大きく回り道をした末に、わたしたちの出発点となったエランという観念、すなわち胚と胚とを結ぶ連結符とも言うべき成体となった有機体を介して、世代から世代へと伝えられていく生命の根源的な弾みという観念に連れ戻されます。諸々の進化系統に分散しながらも存続するこの弾みは、変異の根本的な原因、少なくとも規則的に遺伝し、累積され、新しい種を創造する変異の根本的な原因です。一般に共通の幹から分岐した種は、進化すればするほどより一層分岐する傾向を強めていきます。しかし共通の弾みという仮説が正しいなら、それらの種は幾つかの点では同じように進化し得る筈であり、同じように進化しなければなりません。残された課題は、わたしたちが選んだ例、軟体動物と脊椎動物の眼の形成の例でこのことをより明確に示すことです。それによって「根源的な弾み」とはどういうものか、より一層明らかにすることができるでしょう。
眼のような器官を観察すると、対照的な二つの点、すなわち構造の複雑さと機能の単純さに同時に驚かされます。眼は鞏膜、角膜、網膜、水晶体などそれぞれ異なる部分で構成されていますが、それら各部分もまた、無数の部分に細分することができます。網膜だけを取ってみても、周知のようにそれは神経節細胞、双極細胞、視細胞という三つの神経細胞の層が積み重なったものであり、それぞれ独自の個性を持った各層が極めて複雑な有機体を形成しています。しかし今辛うじて示し得たのは、網膜の精緻な構造の簡略的な図式に過ぎません。眼を機械に喩えると、眼という機械は、それ自体が極めて複雑な無数の機械で構成されている、と言うことができます。一方、見るという行為は一つの単純な事実です。眼を開けば、それだけで視覚が働きます。機能が単純であるだけに、無限に複雑なこの機械を組み立てるに当たって、自然がほんの少しでも手を抜けば視覚は不可能になってしまうに違いありません。器官の複雑さと機能の単純さとのこの際立った対照は、わたしたちの精神を当惑させます。
機械論は、この眼という機械が外的環境の影響の下に次第に形成されてきた、とわたしたちに教えます。外的環境は組織に直接的に作用する場合と、適者を選択することによって間接的に組織に作用する場合があります。しかし機械論がどんな形を取るにせよ、また機械論は、或る程度までは細部の説明に役立つのは事実であるにせよ、眼という機械の部分相互の相関関係についてはわたしたちに何も教えてくれません。
そこで、目的論の出番となります。目的論によれば、機械の諸部分は、あらかじめ立てられた計画に基づき、何らかの目的のために寄せ集められたものだと言います。この点で、目的論は自然の働きを職人の仕事と同一視しています。職人は確かに、或る観念を実現するために、また或るモデルを模倣するために、諸部分を集めることによって事を進めます。それゆえ機械論が、目的論の持つ擬人論的な性格を非難するのは間違いではありません。しかし機械論は、自分自身同じ方法に従っていること、目的論の辿った道筋を、一部を除きそのままなぞっているに過ぎない、ということを見落としています。追求すべき目的とか、手本となるモデルといった観念を機械論が排除したのは事実ですが、自然は職人と同様、諸部分を寄せ集めることによって事を進める、と考える点では機械論も目的論と変わりがありません。胚の成長過程を少しでも観察すれば、生命の作用は、そういう(諸要素の寄せ集めという)やり方とは全く異なっていることにすぐさま気付く筈です。生命は諸要素の結合や累積によって進展するのではなく、分離と分裂によって進展します。
したがって機械論の観点と目的論の観点は、どちらも乗り越えられなければなりません。実を言うと、両者はいずれも、人間の精神(知性)が自分の仕事を省みることによって自ずと導かれた観点に過ぎません。しかしそれらを乗り越えると言っても、どの方向を目指せばよいのでしょうか。先に述べたように、或る器官の構造を分析していくとどこまでも細かく分解できるのに引き換え、その機能は極めて単純です。器官の無限の複雑さと機能の極端な単純さのこの対照こそ、問題解決への糸口となるに違いありません。
一般に、或る対象が一面では単純に見え、他面では無限に多くの要素によって構成されているように見えるとき、これら二つの面は、同じ重要性、と言うより同程度の実在性を持つわけではありません。単純さが対象そのものに属しているとすれば、無限の複雑さは、わたしたちが対象の周囲を回りながらその対象を捉える観点に、別の言い方をすると、わたしたちの感覚や知性がその対象を表象する際に並列する記号に、より一般化して言えば、わたしたちが対象を模倣しようとする際に用いる諸要素に、ただし対象とは秩序の異なる諸要素に属しています。対象はそれらの諸要素とは本性が異なる以上、両者の間に共通の尺度は存在し得ません。絵画の巨匠がカンヴァスに描いた絵を思い浮かべてみましょう。わたしたちは様々な色彩のモザイクの小片を用いて、その絵を模倣することができます。モザイクの小片が小さければ小さいほど、また小片の数が多ければ多いほど、そして小片の色調が多彩であればあるほど、原画の曲線やニュアンスをより正確に再現することができるでしょう。しかしその巨匠が頭の中で単純なものとして思い描き、渾然一体のものとしてカンヴァスに定着させようとしたイメージ、不可分な直観の投影として現れれば現れるほど完璧なものに近付いていくイメージと完全に等しいものを得ようと思えば、無限に小さく、無限のニュアンスを持った無数の要素が必要になります。さてここで、わたしたちの眼は、この巨匠の作品がモザイクの効果としか見えないように作られていると想定してみましょう。或いはわたしたちの知性は、カンヴァスに描かれた絵をモザイクによる創作としてしか理解できないように出来ていると想定してみましょう。その場合、わたしたちは寄せ集められたモザイクの小片の堆積以外のものについて語ることができず、機械論的な仮説に依拠することになります。或いは作品の創作には、寄せ集められた物質的な材料に加えて、モザイク師が仕事を進めるための計画が必要であった、と主張することもできるでしょう。その場合には、わたしたちは目的論の立場を取ることになります。しかしどちらの立場を取るにせよ、わたしたちが創作の真の過程に辿り着くことはありません。何故なら、そもそも寄せ集められた小片などどこにも存在していないからです。存在するのは絵そのもの、つまりカンヴァスに投影された単純な行為であって、わたしたちがそれを知覚したというただそれだけの理由によって、その行為はわたしたちの眼に対して幾千もの小片に分解されます。分解された小片の配列が驚くべきものに見えるのは、小片が再構成されることでその配列が出来上がった、という風に考えてしまうからです。同様に、複雑極まりない構造を持った眼も、見るという単純な行為以外のものではありません。その単純な行為が自ら分解してわたしたちにとって細胞のモザイクとなり、モザイク全体が一つの集合体として表象される結果、その秩序が奇跡的なものに見えてくる、というわけです。
わたしが点Aから点Bまで手を挙げるとき、この運動はわたしにとって同時に二つの形を取って現れます。内からこの運動を感じる場合、それは単純で不可分な行為として現れます。外からこの運動を見る場合、それは一つの曲線ABとして現れます。その曲線上に、わたしは好きなだけ多くの位置を区別することができ、またその曲線そのものも、それらの位置が何らかの秩序に従って相互に配列されたものとして定義することができます。しかし無数の位置にせよ、或いはそれらの位置を相互に結び付ける秩序にせよ、点Aから点Bに手を動かす不可分な行為から自動的に生じたものに過ぎません。機械論がそれらの位置に着目するのに対し、目的論は位置の秩序を考慮に入れます。その反面、機械論と目的論は、実在そのものである運動を揃って見逃してしまいます。運動は、或る意味では諸々の位置やその秩序以上のものです。何故なら運動が与えられさえすれば、それが不可分で単純なものである限り、継起する無数の位置やその秩序も同時に与えられ、それに加えて位置でも秩序でもない本質的な或るもの、すなわち動性が与えられるからです。しかし別の意味では、運動は一連の位置やそれらを結び付ける秩序以下のものです。何故なら諸々の点を或る秩序に従って配列するためには、まずその秩序を表象し、次いで諸々の点でもってこの秩序を実現しなければならないからです。つまり諸々の要素を寄せ集める作業と、知性が必要だからです。ところが現実には、手の単純な運動にはそういったものは全く含まれていません。手の運動は、人間的な意味では知性的なものではありませんし、諸要素から構成されているわけではない以上、それらの寄せ集めでもありません。眼と視覚との関係についても同じことが言えます。視覚には、眼を構成する諸々の細胞や、それら相互の結び付き以上のものがあります。この意味で、機械論も目的論も不十分です。が、別の意味では機械論と目的論は行き過ぎです。何故なら機械論も目的論も、無限に複雑な無数の要素を視覚という単純な行為に仕上げるという、ヘラクレスに命じられたどんな難題よりも難しい仕事を自然に割り当てていますが、自然は眼を作るのに、(ヘラクレスのような怪力どころか)わたしが手を挙げるのと同じ程度の労力しか必要としなかったからです。自然の単純な行為は自動的に無数の要素に分解されるのであって、それらの要素が同じ観念の下に秩序づけられていることにわたしたちは事後的に気付くに過ぎないのです。ちょうどわたしの手の運動が無数の点を(その軌道上に)残していった後で、初めてそれらの点が同じ一つの方程式を満たしていることに気付くように。
しかし以上のことを理解するのは、決して易しいことではありません。というのもわたしたちは、有機的組織化を一種の製作として表象せずにはいられないからです。しかし(そういったわたしたちの先入観に反して)、製作と有機的組織化とは飽くまで別のものです。まず製作に関して言えば、それは人間に固有の活動です。製作とは、材料を様々な部分に分割した上で、部分同士を互いに組み合わせて一つの共通の機能が得られるようにそれらを寄せ集めることです。寄せ集めた諸部分を、わたしたちはその機能の周囲に、すなわち、それら諸部分の言わば中心を占めると想定された機能の周囲に並べていきます。したがって製作は、周辺から中心に、哲学者の表現に従えば多から一に進みます。他方、有機的組織化の働きは中心から周辺へと進みます。それはほとんど数学的な点と言ってよい一つの点から始まり、その点を中心として、徐々に大きさを増していく同心円状の波動によって周辺に拡がっていきます。また、扱う材料が多ければ多いほど効果が増していく製作の働きは、集中と圧縮によって進められます。反対に有機的組織化の作用には爆発を髣髴とさせるものがあります。それは出発点においては、極めて小さい場所と最小限の材料しか必要としません。有機的組織化の力は(本来自己の外に出ることを潔しとせず)、不承不承空間の中に入っていった、という印象を受けます。事実、胚生の過程において原動力となる精子は、有機体の中で最も小さい細胞の一つです。しかも胚生の過程と直接かかわりがあるのは、精子の極く一部でしかありません。
とは言え上述した製作と有機的組織化との相違は、表面的な差異に過ぎません。それらの差異を掘り下げていけば、もっと深い差異が見つかる筈です。
製作されたもの、すなわち製作物は、製作という作業の形式を描いています。ということは、製作者はその製作物のうちに、彼が注ぎ込んだだけのものを再発見することができる、ということです。例えば一つの機械を作ろうとするとき、製作者は機械の部品を素材から一つずつ切り出し、次いでそれらを寄せ集めます。完成した機械のうちには、部品の一つ一つと、部品の総体を見て取ることができるでしょう。ここでは結果の総体が作業の総体を表しており、作業の各部分には結果の或る一部分が対応しています。
実証科学は、有機的組織化を製作と同じ種類の仕事と看做すことができ、またそう看做さなければならないことはわたしも認めます。そうした条件においてのみ、実証科学は有機体の研究に着手することができます。実際、実証科学の目的は事物の基礎を明示することではなく、事物に働きかける最良の手段を提供することです。ところで、物理学と化学は既に学問として確立されています。そしてわたしたちが生ある物質に働きかけることができるのは、それが物理学や化学の方法で扱える場合に限られます。したがって有機的組織化を科学的に研究するためには、有機体をまず機械に見立てなければなりません。そこで細胞は機械の部品と看做され、有機体はそれらの部品の集合体と看做されることになるでしょう。さらに各部分を有機的組織化する部分的な働きは、全体を有機的組織化する働きの実在的な要素と看做されます。こうした観点が科学の観点です。わたしたちの考えでは、哲学の観点はそういったものとは全く異なります。
なるほど、有機的組織化された機械(器官)の全体は有機的組織化の働きの全体を表していると言ってよいかも知れません(と言っても近似的にそう言えるに過ぎません)が、この機械(器官)の各部分は、(有機的組織化という)作業の各部分には対応していません。というのもこの機械(器官)の物質性が表しているのは、製作物の場合とは違い、用いられた手段の全体ではなく、回避された障害の全体だからです。それは肯定的な実在と言うよりも、寧ろ一つの否定です。前著で示したように、理論上、視覚はわたしたちの目の届かない無数の事物にも達し得る能力です。ところがそのような視覚は、行動にまで延長されることは(なく現実のものになることも)ありません。したがってそれ(理論上の視覚)は、生物よりも寧ろ亡霊に相応しい視覚です。生物の視覚は有効なものでなければならず、生物が働きかけ得る対象に範囲が限定されています。生物の視覚とは運河の開鑿のごときものであり、視覚器官は運河を開鑿する作業を象徴しているに過ぎません。運河の開鑿は、両岸に運び出され、積み上げられた土によって説明することはできないように、視覚器官の創造は解剖学的要素の集積によって説明することはできません。機械論者ならば、運河は土を積んだ荷車を一台ずつ運び出して造られたのだ、と説明し、目的論者はそれに付け加えて、土は計画もなく運び出されたのではなく、労働者が一つの計画に従って運び出したのだ、と説明するでしょう。しかし機械論者の説明も目的論者の説明も、どちらも見当違いと言う他はありません。何故なら運河は土(障害物)の運搬作業や積み上げ作業とは別のやり方で造られたものだからです。
わたしたちは先ほど、自然が眼を形成する過程を手を挙げるという単純な動作になぞらえましたが、運河の比喩よりも、手の比喩の方が的確です。ただその際わたしたちは、手は何の抵抗にも出会わないと想定しました。そこで今度は、わたしの手は空気中を動くのではなく、鉄屑の中を進んでいくものとしましょう。わたしが手を前へ進めれば進めるほど鉄屑は圧縮されて抵抗を増していき、或る瞬間が来ると、わたしの手は最早それ以上先へは進めなくなります。ちょうどその瞬間、鉄屑は或る一定の形、すなわち停止した手と腕の一部の形そのままに配列され、秩序づけられます。このとき手と腕が仮に透明であったとすると、それを見ている人は、鉄屑がそのように配列されている理由を(手の動きに求めるのではなく)、鉄屑そのものうちに、或いは鉄屑の堆積に内在する力に求めるに違いありません。或る人は鉄屑一つ一つの位置を、隣接する鉄屑がそれに及ぼす作用と関連付けるでしょう。そういう人は機械論者です。また別の人は、全体的な一つの計画が鉄屑の個々の作用を細部に至るまで支配しているに違いない、と考えるでしょう。そういう人は目的論者です。しかし実際には、そこには単に一つの不可分な行為、手が鉄屑の中を進んでいくという行為があったに過ぎず、鉄屑の筆舌に尽くし難い動きの一つ一つやその最終的な配列の秩序は、この不可分な手の運動を言わば否定的に表現しているに過ぎません。それは抵抗の全体的な形であって、鉄屑の個々の積極的作用の総合ではありません。鉄屑の配列を「結果」と呼び、手の運動を「原因」と呼ぶとすれば、結果の全体は原因の全体によって説明される、とは言い得るでしょうが、原因の各部分に結果の各部分が対応することは決してないでしょう。この場合、機械論の説明も目的論の説明も用をなさず、独自の説明の仕方を見つける必要があります。例えばわたしたちが提示する仮説では、視覚と視覚器官との関係は、おおよそ手と鉄屑との関係に、すなわち手と、手の運動を描き、手の運動によって運河を掘られ、手の運動を限定する鉄屑との関係になぞらえることができます。
手の努力が大きければ大きいほど、手はそれだけ鉄屑の奥まで進むことができます。そして手がどこで止まるにせよ、鉄屑は瞬間的かつ自動的に均整が取れ、相互に調整されます。視覚と視覚器官との関係においても事情は同じです。視覚を構成する不可分な行為がどこまで進むかに応じて、視覚器官の物質性をなす要素、相互に調整された要素が多くなったり少なくなったりします。にもかかわらずその秩序は、どんな場合にも完全無欠です。それが不完全であることは決してありません。何故ならもう一度言いますが、秩序を生み出す実在的な過程には部分が存在しないからです。これこそ、機械論者と目的論者がともに見逃している点であり、わたしたちが眼のような器官の精妙な構造に驚く際に見逃している点です。わたしたちの驚きの奥底には、実現されたのは全体の秩序の一部だけであろう、という思い込み、完全な秩序の実現は一種の恩寵である、という思い込みが常に潜んでいます。目的論者は、この恩寵が目的因によって一度に与えられる、と考えます。機械論者は、この恩寵が自然淘汰によって少しずつ与えられる、と考えます。しかしどちらも秩序のうちに肯定的なものを見ていることに変わりはなく、したがって両者はいずれも、秩序を生み出すその原因のうちに分割可能なもの、あらゆる可能な完成度を許容するものを見ています。実際には、(生命の領域では)原因の強さに程度の違いがあっても、その結果はひと纏まりのものとして、完全な形でしか与えられません。この原因が視覚の方向にどの程度進んでいるかに応じて、下等有機体の単なる色素群が生み出されたり、セルプラ(環形動物)の原基的な眼が生み出されたり、アルキオパ(同じく環形動物)の組織の分化が見て取れる眼が生み出されたり、鳥の驚くほど完成された眼が生み出されたりします。これらの器官は複雑さの程度は違えど、常に同じように調整されています。そういうわけで、二つの異なった動物種の系統がどんなにかけ離れていようが、双方の視覚への歩みが同じ程度に進んでいる限り、どちらの種にも同じような視覚器官が見られるでしょう。何故なら器官の形態は、機能がどの程度発達したかを示す尺度でしかないからです。
しかし視覚への歩みなどという言葉を持ち出すと、目的性という古い考え方に逆戻りすることにはならないでしょうか。もしこの歩みが、意識的なものか無意識的なものかを問わず到達すべき目標を表すものならば、確かにその通りかも知れません。しかし実際には、この歩みは生命の根源的なエランによって行われ、その運動そのものの中に含まれています。だからこそ、この歩みは互いに独立した進化系統の上に見出されるのです。ところでこの歩みは何故、またどのようしてこの運動の中に含まれるのでしょうか。そう問われれば、わたしたちは次のように答えるでしょう。生命とは、何よりもまず生(なま)の物質に働きかける(つまり行動しようとする)一つの傾向である、(だからこそそれはその運動の中に含まれ、そのような傾向としてその運動の中に含まれるのだ)と。この働きの方向は、恐らく前もって決定されているわけではありません。そこから、生命が進化の途上撒き散らしていった形態の予測不可能な多様性が帰結します。この働きは、程度の違いこそあれ常に偶然性を帯びています。或いは偶然性とは言わないまでも、少なくとも選択の兆しを含んでいます。ところで選択は、幾つかの可能な行動をあらかじめ表象することを前提としています。したがって行動そのものに先立って、行動の様々な可能性が生物に素描されていなければなりません。視覚的知覚とはそれ以外のものではありません。目に見える物体の輪郭は、それらに対するわたしたちの可能的行動の素描です。視覚が様々な程度で、極めて多くの動物に見られるのはそのためです。視覚は、それが同じ強度に達するところではどこでも、同じ複雑さを持つ構造を通して姿を現すでしょう。
わたしたちは一般に器官の構造の類似について、その中でも特に眼の構造の類似について論じてきました。というのも一方で機械論に対して、他方で目的論に対して、わたしたちの態度を明確にしなければならなかったからです。次に、わたしたちの態度そのものをより正確に記述しなければなりません。次章において、わたしたちは進化の多岐にわたる成果、ただし今度はそれらの類似している点ではなく、それらが相互に補完し合っている点を検討することによって、わたしたちの考えをより明確にしていきたいと思います。
(つづく)







※コメント投稿者のブログIDはブログ作成者のみに通知されます