









概要
日本の伝統的な大豆発酵食品である納豆は、栄養価が高く、健康に有益であることがよく知られている。
本研究では、納豆が重症急性呼吸器症候群(SARS-CoV-2)や牛ヘルペスウイルス1(BHV-1)などの
ウイルスによる感染を阻害するかどうかを調べた。
興味深いことに、納豆エキスで処理したSARS-CoV-2およびBHV-1は、いずれも細胞への感染を完全に抑制した。
また、BHV-1の糖タンパクDはウェスタンブロット分析で分解されることが示され、
組換えSARS-CoV-2受容体結合ドメイン(RBD)は納豆エキスとインキュベートするとタンパク分解されることがわかった。
さらに、点変異(UK変異体N501Y)を有するRBDタンパクも納豆エキスで分解された。
納豆エキスを100℃で10分間加熱すると、SARS-CoV-2およびBHV-1の細胞への感染能力は回復した。
熱による不活化の結果と一致して、セリンプロテアーゼ阻害剤は納豆エキスによる抗BHV-1活性を阻害した。
したがって、今回の結果は、納豆エキスがウイルスタンパクのタンパク分解を通じて
ウイルス感染を阻害するプロテアーゼを含んでいることを初めて証明するものである。
1.はじめに
重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2)によるパンデミックが終息する兆しはない。
1年以上前に最初の感染が報告されて以来、SARS-CoV-2に関する報告は世界中で劇的に増加している。
最近の技術の進歩により、ワクチンはかつてないスピードで開発されている。
しかし、ワクチンを必要とするすべての人に免疫をつけるには時間がかかる。
さらに、ワクチンの標的エピトープが変異した株も報告されている。
ワクチンだけでは予防できないかもしれない。
したがって、SARS-CoV-2感染に対する他の対策や治療法を開発することが重要である。
本研究の目的は、納豆がSARS-CoV-2および牛ヘルペスウイルス1(BHV-1)を含む抗ウイルス活性を有するかどうかを調べることにある。
BHV-1はウシの呼吸器疾患を引き起こす。納豆は、蒸した大豆を枯草菌で発酵させたもので、
日本で最も伝統的でユニークな食品の一つとして知られている。
伝統的に、日本人は納豆が健康に有益であると信じてきたが、近年、研究によってこの信念を裏付ける科学的根拠が明らかになってきた。
しかし、納豆の抗ウイルス特性は、環状リポペプチド抗生物質であるサーファクチンを除いてはあまり研究されていなかった。
本研究では、納豆エキス中のプロテアーゼ活性がSARS-CoV-2受容体結合ドメイン(RBD)とBHV-1の糖タンパクDを分解し、
細胞へのウイルス感染を阻害することを示した。
2.材料と方法
2.1 納豆エキスの調製
以下に示す方法で納豆エキスを調製した。
市販のS903納豆(タカノフーズ株式会社、茨城県、日本)10gを50mLの遠沈管に入れ、室温で5分間ガラス棒で攪拌した。
その後、リン酸緩衝生理食塩水(PBS)10 mLを加えた。
50回転倒混和した後、混合物を3500 rpmで10分間遠心分離し、上清をφ0.45 μmフィルター、続いてφ0.22 μmフィルターでろ過した。
得られた可溶性画分(以下、納豆エキスと呼ぶ)は、使用するまで-30℃で保存した。
この納豆エキスを、100℃で10分間加熱する処理と、加熱しない処理を行った。
こうして、加熱納豆エキスと非加熱納豆エキスの2種類の試料を調製した。
2.2 ウイルスと細胞
BHV-1石川株(ウシ/日本/1988)はMDBK細胞を用いて培養した。
SARS-CoV-2は、2020年2月に日本のクルーズ船ダイヤモンド・プリンセス号でCOVID-19を発症した患者から分離された株
(SARS-CoV-2/Hu/DP/Kng/19-027、LC528233)を神奈川県衛生研究所から入手した。
SARS-CoV-2は膜貫通型プロテアーゼセリン2(TMPRSS2)を発現するVero E6細胞を用いて培養した。
2.3 プラーク形成アッセイ
上記の2種類の納豆エキス各90μLとBHV-1またはSARS-CoV-2ストック液10μLを混合し、37℃で1時間インキュベートした。
その後、最小必須培地(MEM)で10000倍または5000倍に希釈し、その500μLを12穴または6穴プレートに入れたBHV-1ではMDBK細胞に、
SARS-CoV-2ではVeroE6/TMPRSS2細胞に接種した。
1時間の吸着後、細胞を滅菌PBSで1回洗浄し、カルボキシメチルセルロースを含むMEMで重層した。
37℃で3日間培養した後、感染性を可視化するためにクリスタルバイオレットを用いてプラーク形成アッセイを行った。
基本的に、各試験について少なくとも2回の独立した実験を行った。
試験した納豆エキス単独では、細胞に対する細胞毒性作用は見られなかった(データは示さず)。
2.4 プロテアーゼ阻害剤
PBSで10倍に希釈した納豆エキス80μLを、ハルトプロテアーゼ阻害剤カクテルEDTAフリー(サーモフィッシャーサイエンス株式会社、東京)
1μLおよびPBS 19μLとともに37℃で1時間インキュベートした。
阻害剤カクテルには6種類の阻害剤が含まれている。
アプロチニン(米国ミシガン州ケイマンケミカル)、AEBSF塩酸塩 (AdipoGen Life Sciences, Inc.サンディエゴ、米国)、
ペプスタチン A (Merck KGaA、ダルムシュタット、ドイツ)、ロイペプチン (ペプチド研究所、大阪、日本)、
E-64 (Cayman Chemical)、ベスタチン塩酸塩 ( シグマ アルドリッチ、東京、日本)を用いた。
2.5 ウイルスタンパクの分解解析
2.5.1 BHV-1の糖タンパクD
67.5μLのBHV-1ストック溶液を7.5μLの納豆エキスと37℃で1時間インキュベートした。
抗BHV-1糖タンパクDモノクローナル抗体(1B8-F11 BHV-1/IBR gD-gIV; Veterinary Medical Research and Development, WA, U.S.A)
を用いてウェスタンブロット解析を行った。
2.5.2 SARS-CoV-2の受容体結合ドメイン(RBD)
本研究では、C末端にHisタグを結合した組換えSARS-CoV-2(S1サブユニットタンパク)宿主細胞受容体結合ドメイン(RBD)
(RayBiotech Life, GA, USA)を用いた。
RBDタンパクは、Wuhan-Hu-1単離株(GenBankアクセッション番号QHD43416)に基づいて合成した。
UK variant N501Y RBD-His recombinant protein (Sino Biological, PA, USA)も使用した。
各タイプの納豆エキス4.5μLとSARS-CoV-2 RBDタンパク約2μLの混合物を37℃で1時間インキュベートした。
還元条件下でSDS-PAGEを行い、電気泳動後にゲルをCoomassie Brilliant Blueで染色した。
3.結果と考察
納豆エキスに抗ウイルス活性があるかどうかを調べるため、BHV-1を納豆エキスとインキュベートし、プラーク形成アッセイを行った。
図1Aに示すように、1倍、10倍、20倍希釈の納豆エキスはMDBK細胞へのBHV-1感染を阻害した。
さらに納豆エキスを50倍、75倍、100倍に希釈すると、用量依存的にプラークが形成された。
この結果は、納豆エキスに用量依存的な抗ウイルス活性があることを示している。
興味深いことに、納豆エキスを100℃で10分間加熱すると、このような抗ウイルス活性は消失した(図1B)。
枯草菌が界面活性剤として産生するサーファクチンは、抗ウイルス活性を持つことが知られている。
予想通り、我々の追加実験では、BHV-1を非加熱のサーファクチンとインキュベートした場合、
0.1 mg/mLのサーファクチンがBHV-1の感染を阻害することが示された(データは示さず)。
しかしながら、100℃で10分間加熱したサーファクチンとインキュベートした場合、BHV-1はMDBK細胞に感染し続けた(データは示さず)。
従って、我々の手にある抗ウイルス活性を持つサーファクチンは耐熱性であり、本研究で発見された納豆エキス中の推定阻害因子は耐熱性である。
また、納豆エキスをBHV-1と4℃でインキュベートしたところ、抗BHV-1活性は認められなかった(データは示さず)。
したがって、これらの結果を総合すると、納豆エキスの抗ウイルス活性は、サーファクチンではなく、
プロテアーゼなどの酵素によるものであろうという仮説が導かれた。

図1 BHV-1感染に対する納豆エキスの抑制効果
(A) BHV-1を1倍希釈から100倍希釈までの一連の納豆エキスとインキュベートし、プラーク形成アッセイを行った。
(B) BHV-1を1倍希釈の加熱納豆エキスまたは非加熱納豆エキスとインキュベートした。
その後、プラーク形成アッセイを行い、プラーク数をグラフに示した(平均±SE、n=3)。
我々の仮説を確認するため、納豆エキスをプロテアーゼ阻害剤で処理した。
図2に示すように、納豆エキスをプロテアーゼ阻害剤カクテルで処理したところ、BHV-1は明らかに細胞に感染することができた。
この結果は、納豆エキス中のプロテアーゼが熱感受性阻害因子であることを強く示唆した。
さらに、不可逆的なセリンプロテアーゼ阻害剤であるAEBSF HCl(4-(2-アミノエチル)ベンゼンスルホニルフルオリド塩酸塩)を処理すると、
プラークが観察された(図2)。
この結果から、納豆エキス中のセリンプロテアーゼが抗ウイルス活性の候補の一つであることが明らかになった。

図2 納豆エキスによる抗BHV-1活性に対するプロテアーゼ阻害剤の効果
BHV-1を、プロテアーゼ阻害剤カクテル、アプロチニン、AEBSF HCl、ペプスタチンA、ロイペプチン、E-64、
ベスタチンHClで処理した後、10倍希釈で加熱または非加熱納豆エキスとインキュベートした。
その後、プラーク形成アッセイを行い、プラーク数をグラフに示した(平均±SE、n=3)。
推定されるプロテアーゼがin vitroでBHV-1粒子の表面タンパクを分解するかどうかを調べるために、
抗BHV-1糖タンパクDモノクローナル抗体を用いたウェスタンブロット解析を行った。
糖タンパクDはウイルスのエンベロープと細胞膜の融合に重要な役割を果たしている。
BHV-1を加熱した納豆エキスとインキュベートすると、糖タンパクDは約45-77 kDaのブロードバンドとして現れた(図3、レーン2)。
一方、BHV-1を非加熱納豆エキスとインキュベートすると、糖タンパクDのタンパク分解が誘導された(図3、レーン3)。
糖タンパクD抗体によって検出されたバンドは、レーン3のものと比べて明らかに低値(約30 kDa)であった。
したがって、これらの結果は、推定プロテアーゼが糖タンパクDを分解することによってBHV-1感染を阻害することを示唆している。
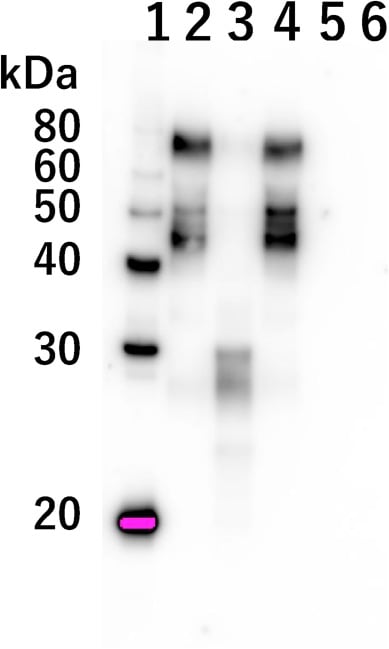
図3. 納豆エキスで処理したBHV-1の糖タンパクDの分解
BHV-1を加熱または非加熱納豆エキスとインキュベートした。
抗糖タンパクDモノクローナル抗体を用いてウェスタンブロット解析を行った。
レーン1:タンパクマーカー
レーン2: PBSで処理したBHV-1
レーン3: 非加熱納豆エキスで処理したBHV-1
レーン4: 加熱納豆エキスで処理したBHV-1
レーン5: 非加熱納豆エキスで処理したPBS
レーン6: 加熱納豆エキスで処理したPBS
次に、納豆エキスがSARS-CoV-2のVero E6細胞への感染を阻害できるかどうかを試験した。
SARS-CoV-2と非加熱納豆エキスを混合した後、プラーク形成アッセイを行った。
図4Aに示すように、SARS-CoV-2を非加熱納豆エキスとインキュベートするとプラークが形成されたが、
加熱納豆エキスではそのような能力は失われた(図4A)。
これらの結果はBHV-1を用いた場合(図1B)と一致しており、納豆エキス中の推定阻害因子は熱感受性プロテアーゼであることが示唆された。
SARS-CoV-2のスパイクタンパクの受容体結合ドメイン(RBD)は、感染初期に宿主細胞のACE2受容体に結合するために重要な役割を果たしている。
図4Bに示すように、33 kDaの組換えRBDタンパクは、非加熱納豆抽出液とのインキュベーションにより明らかに分解された(レーン3)。
一方、加熱納豆エキスを用いた場合、RBDタンパクは無傷であった(レーン4)。
これらの所見を総合すると、図4Aに示した細胞に対するSARS-CoV-2の阻害は、納豆エキスによるRBD分解に起因するという考え方が支持される。
近年、変異型SARS-CoV-2の出現が大きな脅威となっている。
イギリスではRBDのN501Y変異体が優勢になっている。
図4Cは、点変異N501Yを持つRBDタンパクが分解されていることを示している。
さらに、SARS-CoV-2 RBDタンパクの進化的解析から、V367FはACE2受容体への結合親和性を高める重要な変異の一つであることが示された。
V367Yを有するRBDタンパクも納豆エキスで処理することにより分解される(データは示さず)。
これらの結果は、納豆エキスが現在起こっている。
そして、将来起こるであろう多くのタイプの突然変異を分解する可能性を強く示唆している。
図4. 納豆エキスによる抗SARS-CoV-2活性
(A) SARS-CoV-2を加熱および非加熱納豆エキス(1倍希釈)とインキュベートした。
プラークアッセイは3回行った(実験1、2、3)。各プラークアッセイは二重に行った。
実験1と2はパネルAに示した。実験3でも同様の結果が得られた(データは示さず)。
各試験におけるプラーク数は二重測定の平均値である。
(B)SARS-CoV-2の組換えRBDタンパクを加熱または非加熱納豆エキスとインキュベートした。
サンプルはSDS-PAGE用のゲルにロードした。
レーン1:タンパクマーカー
レーン2: PBSで処理したRBDタンパク
レーン3: 非加熱納豆エキスで処理した RBD タンパク
レーン4: 加熱納豆エキスで処理したRBDタンパク
レーン5: 非加熱納豆エキスで処理したPBS
レーン6: 加熱納豆エキスで処理したPBS
(C)点変異(N501Y)を有するSARS-CoV-2の組換えRBDタンパクを加熱または非加熱納豆エキスとインキュベートした。
サンプルはSDS-PAGE用のゲルにロードした。
レーン1: タンパクマーカー
レーン2: PBSで処理したRBDタンパク(N501Y)
レーン3: 非加熱納豆エキスで処理した RBD タンパク(N501Y)
レーン4: 加熱納豆エキスで処理したRBDタンパク(N501Y)
レーン5: 非加熱納豆エキスで処理したPBS
レーン6: 加熱納豆エキスで処理したPBS
本研究では、納豆エキスがBHV-1およびSARS-CoV-2の細胞に対する感染力を阻害し、
BHV-1の糖タンパクDおよびSARS-CoV-2のRBDタンパクを分解することを示した。
しかし、納豆エキスのこの阻害作用は加熱やセリンプロテアーゼ阻害剤で処理すると損なわれた。
この阻害効果は熱に感受性のあるセリンプロテアーゼによるものと考えられる。
このことは、プロテアーゼが糖タンパクDとRBDタンパクの複数の部位を分解することができたという証拠からも裏付けられる。
したがって、納豆に含まれる推定セリンプロテアーゼ(複数可)は、BHV-1およびSARS-CoV-2の感染機能を、
それぞれ糖タンパクDおよびスパイクタンパクのタンパク分解を介して阻害する可能性が示唆された。
今後の研究では、プロテアーゼの同定と切断部位の特定に焦点を当てる必要がある。
納豆エキス中の精製プロテアーゼは、抗ウイルス薬としてさらに研究する価値がある。
最後に、今回の研究結果は、納豆の原料としての新たな応用への道を開くものである。
