獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年5月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ヤンチュアノサウルス・マグヌス

ヤンチュアノサウルス・マグヌスは正しくヤンチュアノサウルス・シャンヨウエンシスの成体である(欧米の研究者の見解)。亜成体であるシャンヨウエンシスに比べてはるかに断片的であるが、2021年にDan Folkes さんが骨格図を出していて、頭が大きめのがっしりした体格になったようだ。PNSOのマグヌスはおおよそそれを立体化したようであり、相変わらず見事な腕前である。確かにヤンチュアノサウルスの顔をしているので購入した。小さい方(シャンヨウエンシス)は今ひとつ気に入らない点があり、まだ買っていない。PNSOの人も他のアーティストの動向をチェックしていることがわかる。
中国の恐竜の中でも四川省は外せない重要な地域である。過去の中国の論文ではヤンチュアノサウルスなどの年代はジュラ紀後期となっていたが、その後ジュラ紀中期とする研究が受け入れられているらしい。ヤンチュアノサウルスーマメンチサウルス類の動物相は、アロサウルスーディプロドクス類や、もっと後のカルカロドントサウルス類ーティタノサウルス類の動物相に先んじて実現していたことになる。長きにわたるアロサウロイドの繁栄の先駆けとなる重要なイベントではないだろうか。
コメント ( 0 ) | Trackback ( 0 )
メトリアカントサウルスはシンラプトルと似ているか
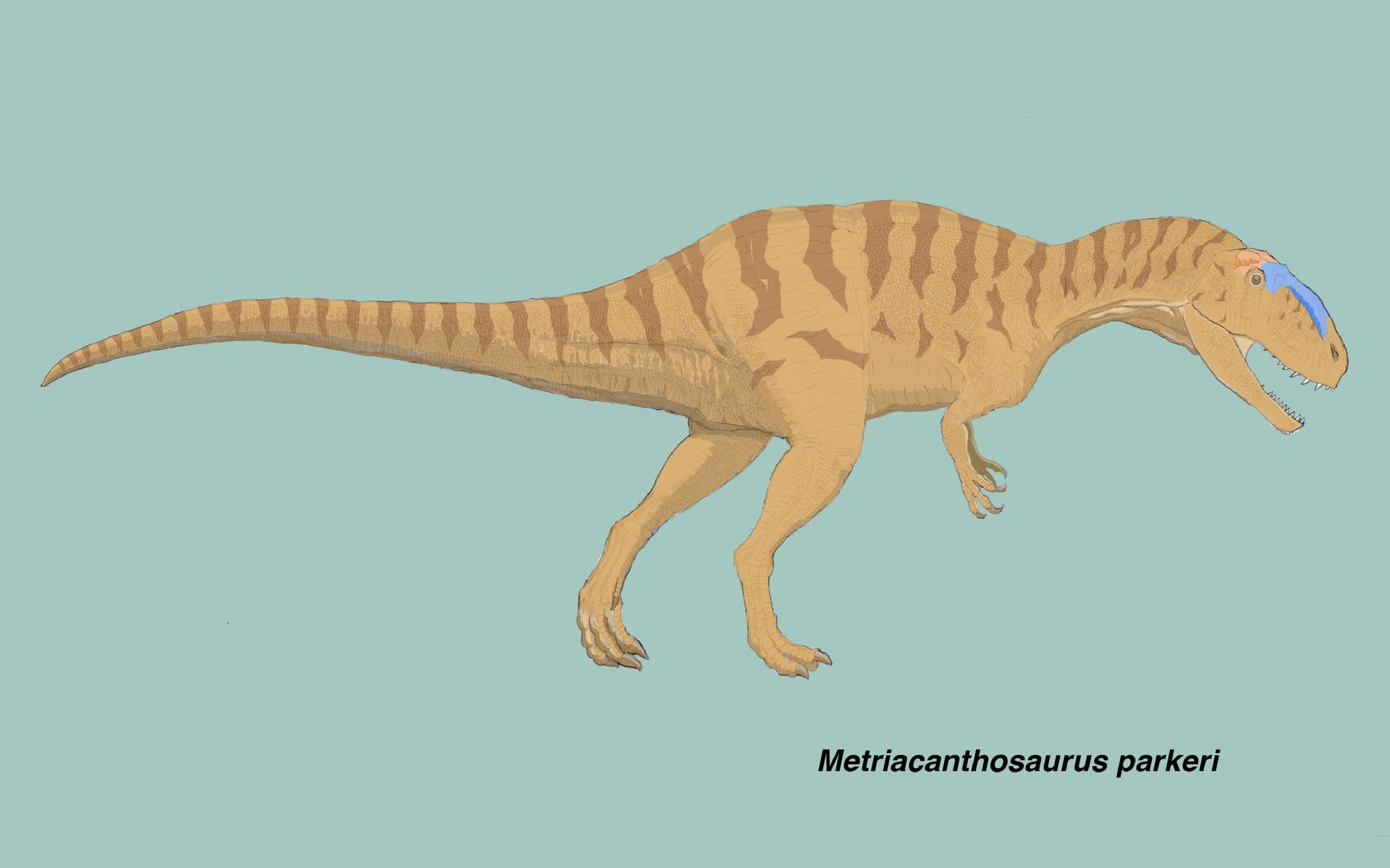
大きい画像
これまで見てきたように、シンラプトルとヤンチュアノサウルスの違いは、腰帯の形質によく表れている。ヤンチュアノサウルスの共有派生形質4つのうち2つは腰帯の形質であり、メトリアカントサウルス亜科の共有派生形質6つのうち3つが腰帯の形質である。そのため、メトリアカントサウルス科の中の分類は、ある程度は、腰帯の形質で読み解くことができる。
それでは、メトリアカントサウルスはどうなのだろうか。メトリアカントサウルスは、常にシンラプトルと共にメトリアカントサウルス亜科に含まれる。腰帯もさぞかしシンラプトルと似ているのだろう、と思って形質データを調べてみた。
メトリアカントサウルスは、胴椎、尾椎、腰帯、大腿骨、部分的な脛骨しか見つかっていない。しかも実は、腰帯の保存状態はよろしくない。断片的な腸骨、恥骨、座骨があるだけである。それでもメトリアカントサウルス科の共有派生形質2つは確かに保持している。298(1) 座骨の断面がハート形(後方にフランジ)と300(1) 座骨の遠位端が癒合している、である。
しかし、メトリアカントサウルス亜科の共有派生形質3つのうち、280(腸骨の後寛骨臼突起の後縁の形)と285(恥骨ブーツの角度)はその部分が保存されておらず、「?」となっている。293(1)(座骨の軸が腹側に曲がっている)は満たしていることになっているが、メトリアカントサウルスの座骨は途中で割れており、遠位の部分はわりと直線的にもみえる。
さらに、ヤンチュアノ側の共有派生形質である281( 恥骨の閉鎖孔などのpuboischiadic plateの状態)は保存されておらず「?」であり、299(座骨の遠位端の形)も「?」となっている。この座骨の遠位端の形は、Walker (1964) の図ではヤンチュアノサウルスのように三角形ではなくシンラプトルと似ているようにみえる。しかしスコアは「?」になっている。
つまりメトリアカントサウルスの腰帯の形質は「?」が多く、メトリアカントサウルス科に含まれるのはいいとして、その中でヤンチュアノサウルスよりもシンラプトルと似ているとは言い切れないようだ。メトリアカントサウルス亜科の共有派生形質6つのうち、腰帯以外の3つは上顎骨、胴椎、脛骨である。メトリアカントサウルスは184(1)(前方の胴椎のキール)と323(1)(脛骨のfibular crestが膨らんでいる)には該当する。総合的にはメトリアカントサウルス亜科に含まれるが、腰帯だけを見ても今ひとつ確認できないということになる。腰帯の図解をしようとしたのに残念である。
当然ながらこの「?」の多さを見ると、メトリアカントサウルスを模式種にしても、他の標本と十分比較できないと感じる。やはり、シンラプトルやヤンチュアノサウルスの仲間という方がわかりやすい。
参考文献
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300. Supplementary material
Weishampel, D. B., Dodson, P. & Osm´olska, H. (1990). The Dinosauria. University of California Press, Berkeley, 733 pp.
コメント ( 0 ) | Trackback ( 0 )
ヤンチュアノサウルスの特徴

麻婆豆腐を狙うヤンチュアノサウルス
科博の「生命大躍進」の時のヤンチュアノサウルス頭骨は、昔の「中国の恐竜展」と同じもので、肉食恐竜好きなら、ほれぼれするような頭骨である。アロサウルス似だが、ちょっとケラト風味が入っている。頭骨の丈が高めで、鼻骨の隆起があり、歯が長めで、下側頭窓が大きい。
ヤンチュアノサウルスは多くのアーティストに描かれてきたが、典型的なカルノサウルス類と書かれていることが多く、具体的な特徴が説明されることは少ないようだ。
最初の記載から40年になるが、実はヤンチュアノサウルスの特徴は定まっていない。
Carrano et al. (2012) によると、ヤンチュアノサウルス・シャンヨウエンシス(Dong et al., 1978)とヤンチュアノサウルス・マグヌス(Dong et al., 1983)の原記載論文に記された特徴は、現在の視点からみるとシンラプトル科、アロサウルス上科、テタヌラ類の特徴を含んでいるという。Currie and Zhao (1994) はシンラプトル科とシンラプトルの特徴は定義しているが、ヤンチュアノサウルスの特徴にはふれていない。
頭骨の高さと長さの比率が、シンラプトルの0.4に対してヤンチュアノサウルスは0.5と、ヤンチュアノサウルスの方が丈が高い。それに伴って上顎骨の丈もやや高い。胴椎の神経棘の高さは椎体の約1.8 倍で、シンラプトルの2.0 倍よりは小さい。またヤンチュアノサウルスでは、頬骨の前眼窩窩の縁が突出しているかもしれないという。しかし詳細な再研究が行われない限り、ヤンチュアノサウルスの特徴を定めることはできないと述べている。
Dong et al. (1983) によるヤンチュアノサウルス・シャンヨウエンシスとヤンチュアノサウルス・マグヌスの違いは、ほとんど大きさの違いに基づいていた。また上顎骨の前眼窩窩の孔の違いは、種内変異とくに成長段階の差による可能性が高く、頸椎の形状の違いは位置による差異とも考えられた。つまりシャンヨウエンシスとマグヌスの形態は事実上同一で、系統解析の形質スコアも同じなので、Carrano et al. (2012) は最初から同一種としている。
原記載が、現在の知見からみると不十分なのは無理もない。1983年といえば、まだシンラプトルもヘピンゲンシスもジゴンゲンシスも、ほとんどのカルカロドントサウルス類もいなかった。他の種類と比較しての考察は、再記載を待つほかない。
ただし昔の論文でも記載自体は生きているはずなので、学ぶ点も多いはずだ。以下は、ヤンチュアノサウルスの文献に書いてあるというよりも、他の文献や筆者の観察を交えながら考えてみたものである。
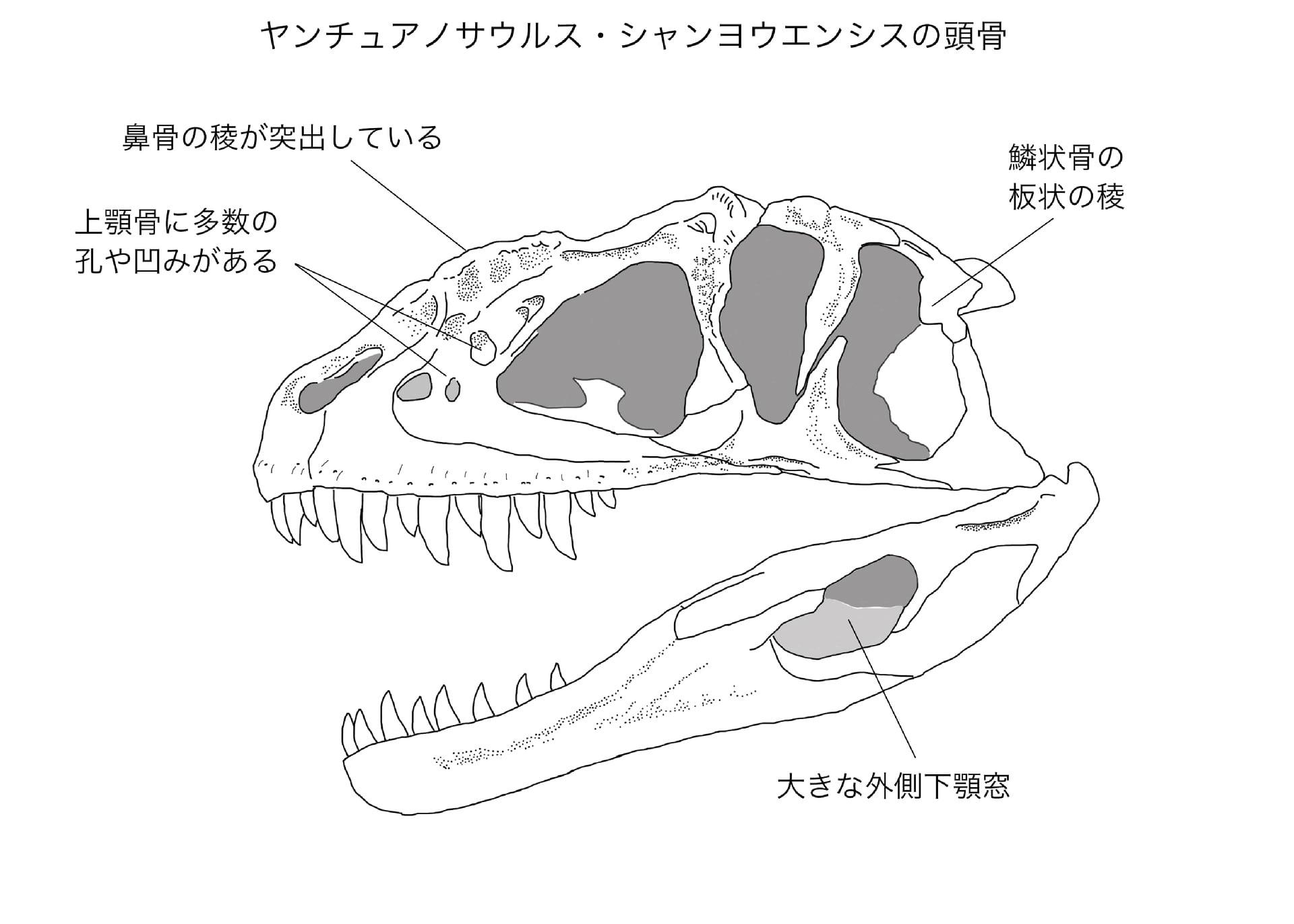
(書き込める情報が少ない。。)
ヤンチュアノサウルスといえば一般的には、鼻骨の側面の稜が大きく発達しているのが特徴のように思われる。実際に他のアロサウロイドには見られないくらい発達していると思うが、意外なことに Dong et al. (1983)の中では、この鼻骨の稜について全く触れていない。ヤンチュアノサウルス属やシャンヨウエンシスの特徴の中にも、鼻骨自体の記載の中にも記述がない。特に注目しなかったのだろうか。Currie and Zhao (1994) は、シンラプトルでは鼻骨の稜が単純で比較的まっすぐであるが、ヤンチュアノサウルスでは鼻骨の稜が強くアーチ状に湾曲し、突出していると述べている。
またシンラプトルでは鼻骨の側面に2個の含気孔がある。ヤンチュアノサウルスの鼻骨でも2個か3個くらい孔があるように見えるが、Dong et al. (1983)には記述されていない。
シンラプトルの記事で述べたように、「シンラプトル科」の特徴として上顎骨の孔がある。シンラプトルと同様にヤンチュアノサウルスでも、上顎骨の前眼窩窩に多数の孔や凹みがある。これについてDong et al. (1983)は、「第1前眼窩窓と第2前眼窩窓(筆者注:maxillary fenestra)の他に、多数の孔や窪みがある」といっているが、詳しい説明はしていない。シンラプトルでは、maxillary fenestra のすぐ後方に1個の孔があり、その後背方に大きな楕円形の窪みがある。その窪みの中には1個の大きい孔と、それより小さい窪みが2個くらいある。ヤンチュアノサウルスの頭骨を観察すると、やはり孔や窪みの配列パターンはシンラプトルとよく似ているように見える。
Dong et al. (1983)はヤンチュアノサウルス属の特徴として、前頭骨と頭頂骨が癒合している(fused)といっている。前頭骨と頭頂骨が癒合しているのはカルカロドントサウルス類やアベリサウルス類にみられる形質であるが、本文中でtightly associated といっているので、co-ossification という意味ではないのかもしれない。
ヤンチュアノサウルスでは、アロサウルスよりも頭頂骨の突起が大きく発達して背方に伸びているとある。しかし他の獣脚類と比べてそれほど目立つわけでもないような気もする。マジュンガサウルスのように目立つわけではないようだ。
Dong et al. (1983)のスケッチでは、後眼窩骨の後方突起(鱗状骨突起)がかなり長いように描かれているが、頭骨レプリカを観察すると、この突起はシンラプトルほどではないが、ある程度短いように見える。この辺りはシンラプトルとの類縁を感じる。
頭骨レプリカを観察していて気がついたが、鱗状骨の形がシンラプトルとは異なるようである。これはDong et al. (1983)の中で記載されている。鱗状骨の前方突起と腹方突起の間に、板状の稜laminar ridgeがあると書いてある。ヤンチュアノサウルスに固有というわけではないだろうが、シンラプトルとは異なる特徴には違いない。
アロサウルスとは異なり、シンラプトルと同様に、下顎には大きな外側下顎窓がある。これはアロサウロイド全体の中でどうなのかわからないが、アベリサウルス類のような原始的な状態なのだろうか。
ヤンチュアノサウルスの歯は一般的な獣脚類の形状をしており、前上顎骨歯4、上顎骨歯14-15、歯骨歯14-15 である。頭骨の左側の歯列は完全に保存されている。左の上顎骨では2, 5, 7, 9 番目の歯が最も大きく、剣状で側扁しており、前縁と後縁に鋸歯がある。これらの歯冠の高さは5.6 cm で後方に反っている。1, 3 番目の歯は比較的小さく、高さが3.5 cm である。11-15 番目は小さく、薄く、最大2 cm であるという。
上顎と下顎が強く閉じているため正確な測定や記載はできないが、歯骨歯についても記述している。
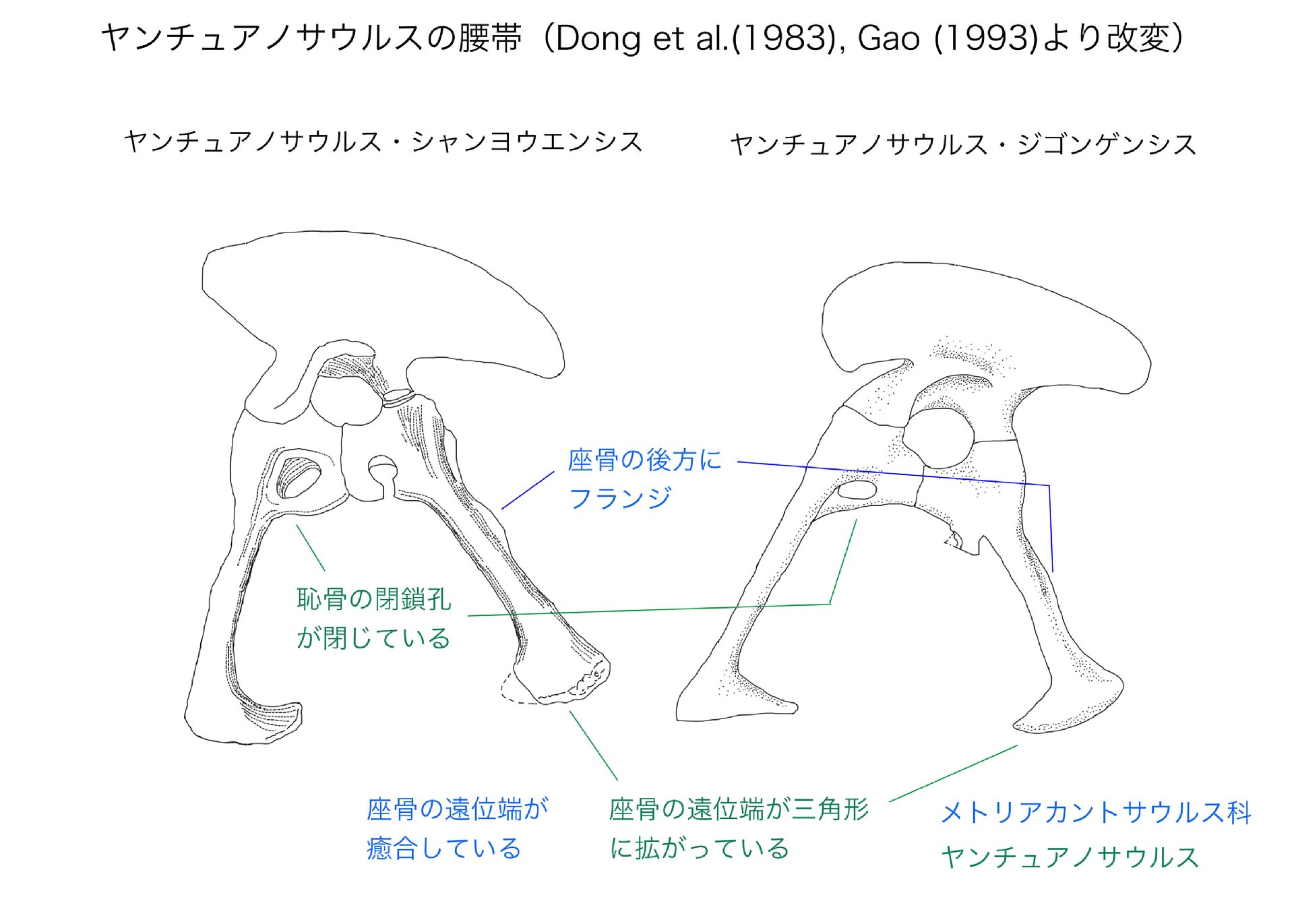
特徴といっても色々あって、メトリアカントサウルス科の中のグループ、つまりクレードとしてのヤンチュアノサウルス属の共有派生形質なら、Carrano et al. (2012) のAppendixを見ればわかる。
ヤンチュアノサウルス属の共有派生形質は4つあり、そのうち2つは腰帯の形質である。1つは座骨の遠位端が三角形に拡がっていることで、ジゴンゲンシスでは特に顕著である。もう一つは恥骨の閉鎖孔という孔が閉じていることである。これは厳密には「恥骨の閉鎖孔が閉じていて、座骨の孔は開いている状態」に相当する。恥骨の基部から座骨の基部にかけて、原始状態では3つの孔があるという。恥骨の閉鎖孔はヤンチュアノサウルスでは閉じているが、シンラプトルやシャモティラヌスでは微妙に開いている。(ちょっと気になるのは、Gao (1993) のジゴンゲンシスの腰帯のスケッチではこの孔が明らかに閉じているが、恥骨の写真を見るとどうも破損しているようにみえることである。この肝心な部分が推定ということはないのだろうか。閉じていると考えて記載しているのは確かなので、何か根拠があるのかもしれない。)
その他、腸骨の後端の形や、恥骨ブーツの角度なども、シンラプトルとは異なることがわかる。このように腰帯を並べてみると、“スゼチュアノサウルス”がなぜヤンチュアノサウルスとまとめられたのか、理解しやすい。
参考文献
Dong, Z., Chang, Y. H.,Li, X. M. & Zhou, S.W. (1978). [Note on the new carnosaur (Yangchuanosaurus shangyouensis gen. et sp. nov.) from the Jurassic of Yangchuan District, Szechuan Province.] Kexue Tongbao, 5, 302-304 [In Chinese].
Dong, Z., Zhou, S. W. & Zhang, H. (1983). [Dinosaurs from the Jurassic of Sichuan.] Palaeontologica Sinica, New Series C, 23 (Whole Number 162), 1-136 [In Chinese].
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300.
コメント ( 0 ) | Trackback ( 0 )
不遇なヤンチュアノサウルス
YouTubeには、Jurassic World Evolution というゲームの、メトリアカントサウルスのCG動画が上がっている。当然ながらほぼシンラプトルであるが、なかなか良いものである。そのコメント欄に、英国の恐竜なので感慨深い、という声があった。なるほどやはり、バリオニクスやメトリアカントサウルスが登場するのは、イギリスの恐竜ファンを意識したものなのだろう。
私は中国人ではないが、なんとなく「シンラプトル科」がアジアで独自に進化した固有のグループだった方が良かった。イギリスのメトリアカントサウルスさえいなければ、「シンラプトル科」のままであった。もちろん古生物地理学上、メトリアカントサウルス類が広くユーラシアに分布していたことは意義のある発見である。
メトリアカントサウルスは断片的な化石であるが、歴史的に早く記載された方が優先権がある。それはわかる。アベリサウルスよりもカルノタウルスの方が完全であるが、アベリサウルス科になったわけである。
しかし、それならばシンラプトル科の樹立の時点で、なぜ「ヤンチュアノサウルス科」にならなかったのだろうか。ヤンチュアノサウルスは1978年と1983年で、シンラプトルの1993(1994) 年よりも先に記載されている。そして、Currie and Zhao は最初からシンラプトル科にヤンチュアノサウルスが含まれるといっている。それなら普通は、「ヤンチュアノサウルス科」になるはずである。
確かにCurrie and Zhaoのシンラプトルの詳細な記載は、この時代の恐竜研究の1つの金字塔のような仕事のようだ。それに比べるとヤンチュアノサウルスの記載は、他の種類との比較が不十分で、標徴形質も不完全なものかもしれない。当時としても正式な記載とはみなされなかったのだろうか?しかし1978年は短報であるが、1983年は正式な記載論文の形をとって出版されているように見える。個々の骨について、簡潔かもしれないが一応は記載している。なぜこれが無視されなければならなかったのか。欧米の研究者には読めない中国語の論文だったからか?
ともあれシンラプトルやヤンチュアノサウルスなどはシンラプトル科になり、主として中国の恐竜として長年認識されてきた。それが今度は、断片的なメトリアカントサウルスに科の名称を奪われた形になってしまった。中国の恐竜ファンは残念ではないのだろうか。
これまたどうでもいいことだが、ヤンチュアノサウルスは昔サファリ社のフィギュアとして商品化されたことがある。しかしそのポーズが、なぜかお辞儀をするように頭を下げた「反省」姿勢だった。もっと普通のポーズにすれば売れたのではないだろうか。私は、閉館間近のユネスコ村恐竜探検館に行った時、このヤンチュアノサウルスが大量に在庫処分されていたのを見て、心を痛めたことがある。
博物館や恐竜展のショップで販売されているカロラータのジュラ紀恐竜セットでも、どうもアロサウルスが主役で、ヤンチュアノサウルスは力が入っていないような気がする。何かと不遇な恐竜ではないか。
コメント ( 0 ) | Trackback ( 0 )
Wikipediaの記述について
ウィキペディアを100%信じる人もいないだろうが、参考にはなるものである。恐竜に関しては通常、日本語版よりも英語版の方が信頼でき、非常に適切な記事も多い。そもそも情報量が違う。日本語のヤンチュアノサウルスの記事は現在、5行ほどしかなく、参考文献がポケット図鑑などという状態である。英語版ははるかに充実していて、複数の原著論文を引用している。
ところが、5月26日現在、英語版のWikipedia でYangchuanosaurusの記事を見ると、ジュラ紀中期と書いてある。しかもマメンチサウルスやオメイサウルスと同時代とある。なぜそうなっているのか不明である。他には変なところはない。
Carrano et al. (2012) を引用して内容をしっかり説明してあるので、現在ではジュラ紀中期から後期というべきだろう。ヤンチュアノサウルス・ジゴンゲンシスだけがジュラ紀中期 (Xiashaximiao Formation)で、シャンヨウエンシス(マグヌス、CV00214含む)はジュラ紀後期(Shangshaximiao Formation)のはずである。Shangshaximiao Formationは、 Carrano et al. (2012) によるとオックスフォーディアンから初期キンメリッジアンとある。
ちなみに、Metriacanthosauridae の記事にはメトリアカントサウリダエの共有派生形質が書いてあるが、12個のうち1個抜けている他に、2箇所誤りがある。arm+forearmのところと、ilium ではなくischium の断面である。メトリアカントサウリナエの中にも1箇所ある。
このように、専門的な記述で埋まっていると誰もチェックしなくなり、誤りがそのままになっていることは問題である。
文章を書く立場としても、注意しなければならないと考えさせられた。
コメント ( 0 ) | Trackback ( 0 )
ヤンチュアノサウルス・ジゴンゲンシス(スゼチュアノサウルス・ジゴンゲンシス)
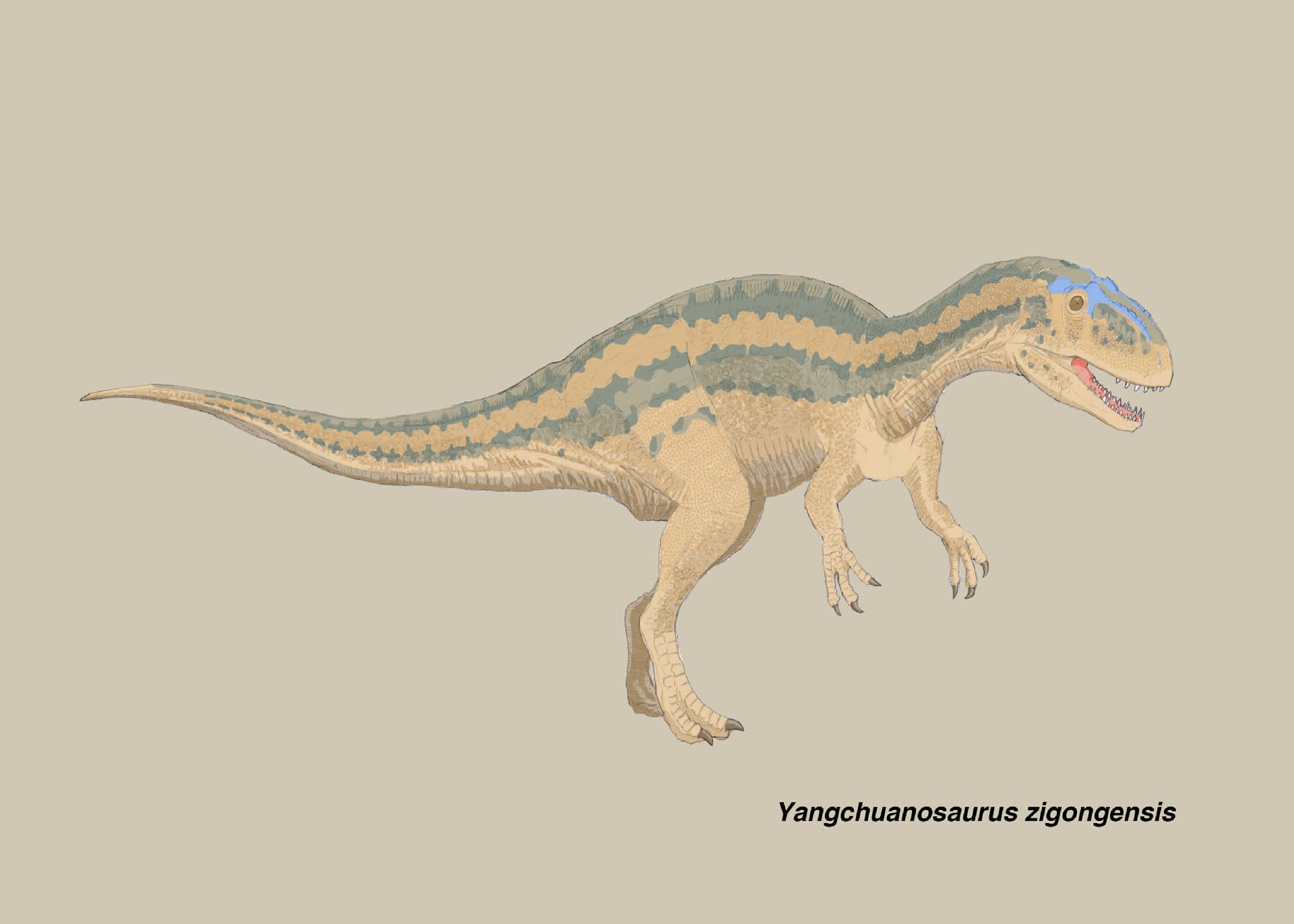
大きい画像
ヤンチュアノサウルスやシンラプトルに比べて、具体的なイメージがつかめないのがスゼチュアノサウルスである。中国の恐竜展などで復元骨格を見るたびに、どこを観察すればいいのか戸惑ってしまう。しかもある意味残念なことに、Carrano et al. (2012) の系統研究で、この「四川竜」の属名は失われてしまった。
スゼチュアノサウルスの歴史
歴史的経緯については、Wikipedia(英語)等にも書いてあるので簡潔にメモする。
Young (1942)が記載したスゼチュアノサウルス・キャンピの模式標本は、4つの分離した歯化石で、同定できる特徴がないために、現在は疑問名とされている。そのため新たに発見された化石にスゼチュアノサウルスの属名を用いることは不適切である。
四川省自貢のジュラ紀後期の地層から発見された部分骨格CV00214は、1978年にスゼチュアノサウルス・ヤンドネンシスとされ、1983年にスゼチュアノサウルス・キャンピとされたが、上述の理由で長年疑問視されていた。頭骨はなく、歯は含まれていない。
さらにGao (1993)は、四川省自貢のジュラ紀中期の地層から発見された部分骨格に、スゼチュアノサウルス・ジゴンゲンシスと命名した。Chure (2001) は、CV00214とスゼチュアノサウルス・ジゴンゲンシスを同一種として、(スゼチュアノサウルスは使えないので)新しく命名すべきであると考えた。
Carrano et al. (2012) の系統解析の結果、CV00214はヤンチュアノサウルス・シャンヨウエンシスと姉妹群となり、それらとスゼチュアノサウルス・ジゴンゲンシスが姉妹群となった。つまり系統関係上も“スゼチュアノサウルス”は解体された。この結果を受けてCarrano et al. (2012)は、同じ地層から出ていることからもCV00214はヤンチュアノサウルス・シャンヨウエンシスと同一とし、より古い地層から出ているスゼチュアノサウルス・ジゴンゲンシスは別種ヤンチュアノサウルス・ジゴンゲンシスとした。新しい属名が必要になったがヤンチュアノサウルス属の別種とすることで系統関係の近さを表したといっている。
ここで気になる点が2つある。1つは、種レベルの違いか属レベルの違いかの判断はやはり、研究者によって異なることである。スゼチュアノサウルスの代わりに、新しい属名がついた方がアマチュア的には楽しかった。
もう1つは、中国四川省は獣脚類化石の宝庫であるが、あまり欧米の研究者がアクセスできていないようにみえることである。Carrano et al. (2012) は、系統解析に用いた標本の90%は、3人の共著者の誰かが直接観察していると述べている。First handで見た種類は太字で表記している。しかし四川省のヤンチュアノサウルスやスゼチュアノサウルスの標本は、直接見ておらず、文献の記述と写真・図版からデータをとっているようである。四川省の研究者が認めているかどうか知らないが、これでは納得しないかもしれない。本当は、中国側と欧米側が共同研究して、結論を出してほしいところである。
ジゴンゲンシスの特徴
ヤンチュアノサウルス・ジゴンゲンシス(スゼチュアノサウルス・ジゴンゲンシス)は、中国四川省自貢市大山舗のジュラ紀中期の地層から発見されたメトリアカントサウルス類で、Gao (1993)によって記載された。自貢恐竜博物館の建設工事中に発見された大量の恐竜化石の1つで、推定6mの肉食恐竜である。
ホロタイプZDM9011は胴体のかなりの部分を含む部分骨格で、連続した完全な10個の頸椎、13個の胴椎、5個の仙椎、不連続な25個の尾椎、左の上腕骨、尺骨、橈骨、橈側骨、中手骨、指骨、右の肩甲骨、完全な腰帯からなる。その他に同時に発見された参照標本としてZDM9012:左上顎骨、ZDM9013:10本の分離した歯、ZDM9014:右の大腿骨、脛骨、腓骨がある。
Gao (1993)はジゴンゲンシスの特徴として、前部・中央部の頸椎は後凹型、後部の頸椎は平凹型で腹側にキールがある、後部尾椎の前関節突起が長い、上腕骨の三角筋稜がよく発達している、橈骨/上腕骨の比率が56%、第IV中手骨が残存している、腸骨は特に丈が低い、恥骨ブーツが小さい、座骨の遠位端が拡がっている、などをあげている。(中国語と英語で内容が微妙に異なる。)
一方Carrano et al. (2012)は、頸椎の特徴だけをあげている。Holtz et al. (2004) に従い、前方の頸椎2-4 だけが後凹型、残りは両平面型であるテタヌラ類としている。後方の頸椎の関節面が平面的ということだろう。Gao (1993)のあげた特徴のいくつかは、メトリアカントサウルス科やヤンチュアノサウルスの特徴に含まれた。
Gao (1993)は、スゼチュアノサウルス・キャンピとされたCV00214とジゴンゲンシスの違いについて記している。ジゴンゲンシスでは、後部頸椎が平凹型で腹側にキールがある;三角筋稜が特に発達している;第IV中手骨が存在する。一方CV00214は、頸椎が後凹型でキールがない;三角筋稜があまり発達していない;第IV中手骨がない。
ヤンチュアノサウルス・シャンヨウエンシスのホロタイプでは前肢は保存されていないが、CV00214がこれと同一種とすれば、完全に3本指だったことになる。
なぜスゼチュアノサウルスがヤンチュアノサウルスになったのかについては、次の記事に持ち越すことにする。
参考文献
Gao, Y. (1993). [A new species of Szechuanosaurus from the Middle Jurassic of Dashanpu, Zigong, Sichuan]. Vertebrata PalAsiatica, 31, 308-314 [In Chinese].
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300.
Shangshaximiao Formation Xiashaximiao Formation
コメント ( 0 ) | Trackback ( 0 )
シンラプトル2018 (2)
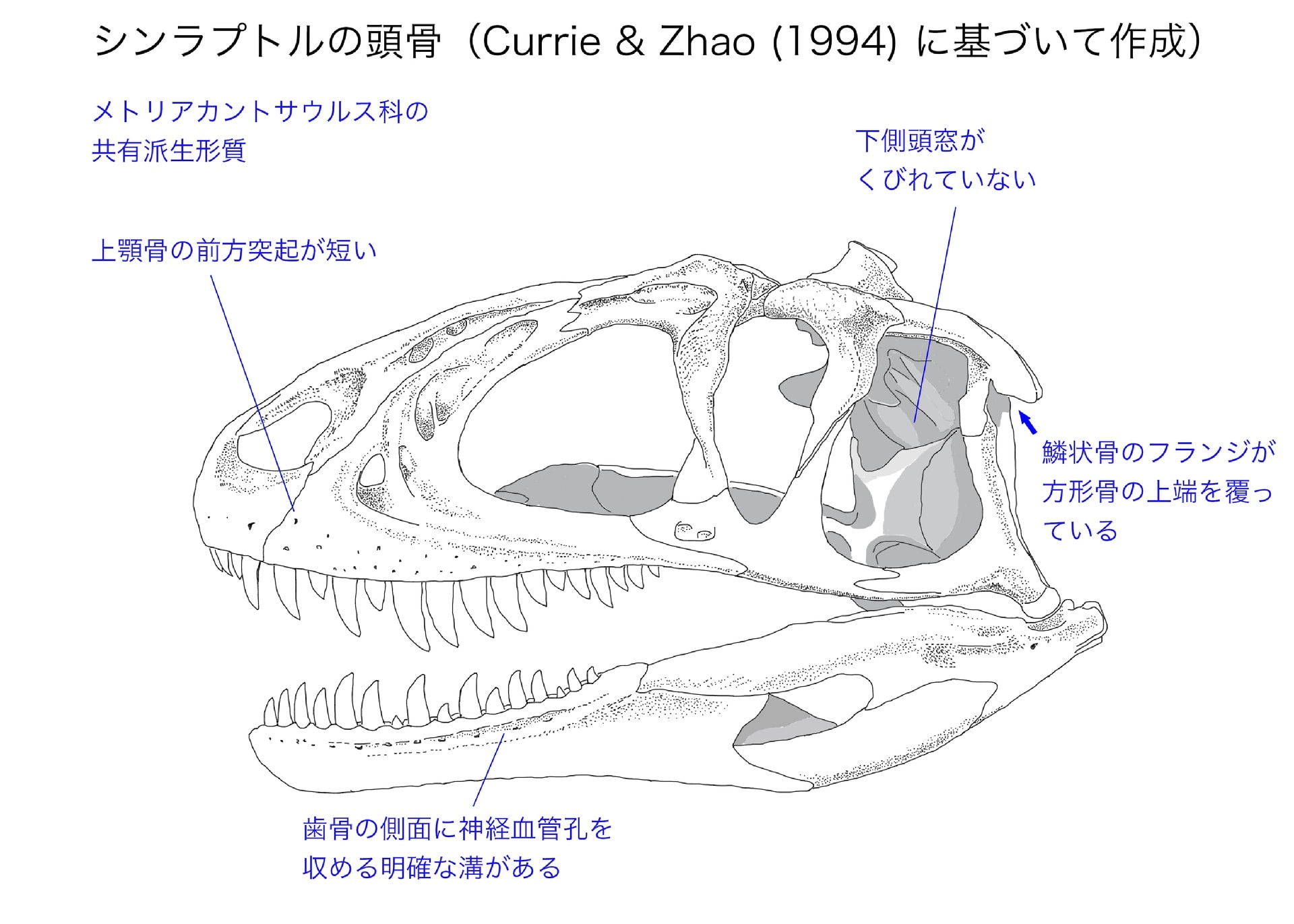
メトリアカントサウルス科の共有派生形質
メトリアカントサウルス科は12の明確な共有派生形質でまとめられるが、細かい形質が多く難解なものもある。その中でわかりやすいのは、前述した第IV中手骨である。「第IV中手骨が存在し、第IV指の指骨と第V指が失われている」に該当する。アロサウルス上科の中では主にメトリアカントサウルス科だけにみられる形質で、シンラプトル、ヤンチュアノサウルス・ジゴンゲンシス、シュワンハノサウルスで確認される。

また少し難しそうだが重要な形質として、軸椎の間椎心intercentrumの向きがある。(頸椎のうち第1頸椎を環椎、第2頸椎を軸椎という。それぞれ本来は間椎心と椎心(椎体)からなっていて…という環椎・軸椎の構造については、人体解剖学や比較解剖学の本に載っている。)
シンラプトルでは歯状突起odontoidは分離して見つかっている。軸椎の腹側では間椎心と椎心(椎体)が結合している。多くの獣脚類(ディロフォサウルス、ケラトサウルス、カルノタウルス、ピアトニツキサウルス、アロサウルス)では軸椎の間椎心の腹側面が水平であるが、シンラプトルでは前背方に傾いている。Currie & Zhao (1994) は、ジュラ紀の獣脚類では他にヤンチュアノサウルスやモノロフォサウルスなど2,3の種にしかみられないと述べている。Carrano et al. (2012) によると、この形質はマーショサウルスを含むいくつかの基盤的なテタヌラ類にみられるという。いずれにしてもメトリアカントサウルス科は「軸椎の間椎心の腹側面が前背方に傾いている」を共有する。
この軸椎の形態は、頸椎を後頭顆の下に寄せることで頭骨をよく支える役割があると考えられている。同じような適応はティラノサウルスやタルボサウルスにもみられるといっている。つまりこれは原始的な形質ではなく、大きな頭骨を支えるための先進的な形質ともいえるのではないか。シンラプトルも頑張っている。

メトリアカントサウルスやシャモティラヌスが近縁とされたということは、腰帯の形質が効いているにちがいない。シャモティラヌスはどこがシンラプトルと似ているのか。
メトリアカントサウルス科の共有派生形質の一つは座骨の形態で、「左右一対の座骨軸の断面がハート形」、つまり正中部分がフランジ状に後方に突出していることである。Currie & Zhao (1994) は座骨の後背方にcrestがあるといっていて、同じくらい発達しているのはヤンチュアノサウルスだけであると述べている。もう一つは「座骨の遠位端が癒合している」ことである。これはシャモティラヌスでは確認できない。
また腸骨の後寛骨臼突起の後縁が直線的であることと、恥骨ブーツが強く後背方に傾いている(恥骨軸と恥骨ブーツの角度が60度より小さい)ことは、メトリアカントサウルス亜科の共有派生形質となっている。
メトリアカントサウルス、シンラプトル、ヤンチュアノサウルスはいずれも胴椎の神経棘が中程度に長いが、そのことはメトリアカントサウルス科の確実な共有派生形質には含まれていない。(ACCTRANという方式の場合のみ入る。)メトリアカントサウルス科の中にはヤンチュアノサウルス・ジゴンゲンシスのように胴椎の神経棘があまり長くないものもいるので、共通祖先の形質ではなく、一部の種類が獲得した形質ということだろう。これもアクロカントサウルスのようなカルカロドントサウルス類との収斂的な傾向かもしれない。
シンラプトルのホロタイプは推定全長7.2 m であるが、脳函の骨同士、脊椎の神経弓と椎体、個々の仙椎同士などが完全に癒合していないことから、亜成体と考えられている。成体はもう少し大型だったかもしれない。全長7.2 mに対して頭骨は90 cm と頭がかなり大きい。中足骨はアロサウルスよりも長い。ただし亜成体であることを考えると、成体はアロサウルスと同じくらいの可能性があるという。
コメント ( 0 ) | Trackback ( 0 )
シンラプトル2018(1)

大きい画像
シンラプトル科からメトリアカントサウルス科へ
シンラプトルやヤンチュアノサウルスの仲間は伝統的にシンラプトル科Sinraptoridaeと呼ばれてきたが、Carrano et al. (2012) のテタヌラ類の系統解析の結果、歴史的に古いメトリアカントサウルスがシンラプトル科に含まれたため、メトリアカントサウルス科Metriacanthosauridaeと呼ばれるようになった。ただし、新しく含まれたメトリアカントサウルスやシャモティラヌスなどは化石が断片的で、明確なイメージがない。依然としてこのグループの代表は中国のシンラプトルやヤンチュアノサウルスであり、はるかに完全な全身骨格による明確なイメージがある。また何度も日本の恐竜展などで公開されているので、昔からの日本の恐竜ファンにとっては、シンラプトル科の方が馴染みがあるだろう。ただし最近の書籍ではメトリアカントサウルス類になっているはずなので、シンラプトル科を知っているのはオッサンの証拠になるかもしれない。
実際には、「メトリアカントサウルスがシンラプトル科に含まれた」という表現は正確でないかもしれない。分岐分析の結果、昔のシンラプトル科のメンバーを含む、拡張されたクレードが出現したということで、とらえ方が異なるのだろう。
メトリアカントサウルス科には、シュワンハノサウルス、シダイサウルス、ヤンチュアノサウルス・シャンヨウエンシス、“スゼチュアノサウルス・キャンピ”とされたCV00214、“スゼチュアノサウルス”・ジゴンゲンシス、メトリアカントサウルス、シンラプトル・ドンギ、シンラプトル・ヘピンゲンシス、シャモティラヌスが含まれた。ただし断片的なシュワンハノサウルスを除外した方がずっと分岐図が安定し解像度も上がった。分岐図はネット上にある。
メトリアカントサウルス科の中には2つのクレード、ヤンチュアノサウルスの仲間とシンラプトルの仲間が含まれる。ヤンチュアノサウルスの仲間は、“スゼチュアノサウルス”の使用が無効とされ、全てヤンチュアノサウルス属となった。一方、シンラプトルの仲間はメトリアカントサウルス亜科Metriacanthosaurinaeとされ、メトリアカントサウルス、シンラプトル・ドンギ、シンラプトル・ヘピンゲンシス、シャモティラヌスが含まれている。
シンラプトル科は、シンラプトルを詳細に記載したCurrie & Zhao (1994) によって樹立された。そこでは、シンラプトルとヤンチュアノサウルスがシンラプトル科に含まれるとしている。しかしその当時は、多くのカルカロドントサウルス類の標本がほとんど発見されていなかったので、アロサウルスやケラトサウルスなど、限られた種類としか比較されていない。そのため当時のシンラプトル科の特徴は変わってくると思われる。例えばシンラプトル科の特徴として、上顎骨の前眼窩窩に2個以上の副次的な孔があることをあげている。現在では、アクロカントサウルスやエオカルカリアにも副次的な孔があることは知られているが、シンラプトルほど多くはないようにもみえる。この特徴の位置づけはどうなったのだろうか。(メトリアカントサウルス科の特徴ではない。)一方、シンラプトル科の特徴のうち軸椎の間椎心の傾きについては、メトリアカントサウルス科の特徴として受け継がれている。

シンラプトルの特徴
Currie & Zhao (1994)ではシンラプトル属の特徴として、頭骨がヤンチュアノサウルスよりも長く、丈が低いこと、頬骨に大きな含気孔があること、後眼窩骨に粗面が発達していることなどが含まれていた。Carrano et al. (2012) では、これらの一部(以下の1)2)3))をシンラプトル・ドンギの特徴として採用している。
Carrano et al. (2012) によるシンラプトル・ドンギの特徴は、アロサウロイドのうちで1)下側頭窓が大きく、比較的長くまっすぐな後眼窩骨-鱗状骨バー、2)後眼窩骨の後方突起(鱗状骨突起)が非常に短くみえる、3)口蓋骨の内部に深く含気性が発達している、の3つである。後眼窩骨-鱗状骨バーとは、上側頭窓と下側頭窓の間を仕切る部分でCurrie & Zhao (1994)ではintertemporal bar と呼んでいたが、Carrano et al. (2012)ではpostorbital-squamosal bar と言い換えている。またCurrie & Zhao (1994)によると、後眼窩骨の後方突起は非常に短くみえるが、内側から見るともっと後方まで伸びていて、外側が鱗状骨に覆われているのであるという。そのためCarrano et al. (2012)は短いと言い切っているが、ここでは「短くみえる」としておいた。
原始的なアロサウロイド
シンラプトルはよく、「アロサウルスよりも原始的」と書かれる。どこが原始的なのだろうか。
Currie & Zhao (1994)では、シンラプトルがアロサウルス科よりも原始的な点として、下側頭窓が大きいことから始まって、距骨の上行突起が比較的低いことまで、非常に多くの形質をあげている。
おそらく最もわかりやすいのは第IV中手骨が残っていることで、テタヌラ類は3本指であるが第4指の痕跡があるということである。これはシンラプトル科の特徴には含まれていなかったが、メトリアカントサウルス科の特徴として含まれている(後述)。
またアロサウルス上科の系統進化を論じる研究では必ず言及されるのが、歯間板の状態である。シンラプトルでは、ドゥリアヴェナトル、ピアトニツキサウルス、モノロフォサウルスのような基盤的なテタヌラ類と同様に、歯間板が分離しており、癒合していない。アロサウルス上科の中で、アロサウルス以上の種類では歯間板が互いに癒合している。さらに派生的なカルカロドントサウルス類では、歯間板の丈が高くなっている。メガロサウルス類の中でも、トルボサウルスなどでは歯間板が癒合し、強化されている。一方小型のコエルロサウルス類出身のティラノサウルス類では、歯間板は癒合していない。つまりティラノサウルス類では顎や歯自体は非常に頑丈になっているが、歯間板の強化については原始的な状態である。
シンラプトルの肩甲骨は、太いメガロサウルスと細長いアロサウルスの中間的な形で、遠位端がはっきり拡がっていない点でアロサウルスよりも原始的とされている。
シンラプトルでは頸椎と前方の胴椎の含気腔が単純で、多数の骨梁trabeculae で仕切られていないとある。これもアロサウルス上科全体の進化傾向からみると、カルカロドントサウリアでは頸椎と胴椎の含気性が発達していくので、シンラプトルは原始的ということが納得できる。
シンラプトルがカルカロドントサウルス類と似ている点
まだアロサウルス上科の系統関係が定まっていない時期、例えばアクロカントサウルスやネオヴェナトルがアロサウルス科とされた頃には、シンラプトルがカルカロドントサウルス類と近縁と考えられたことがある。これは主に脳函のためである。シンラプトルの脳函の形態はカルカロドントサウルス類と似ている点が多く、Coria & Currie (2002) によるとCurrie & Carpenter (2000) のデータに脳函の形質を加えて系統解析すると、ギガノトサウルス、カルカロドントサウルスが姉妹群となりシンラプトルを加えた3種がクレードとなった。この3種には5つの共有派生形質があるとしていた。
最も顕著なのは後頭部の角度で、CTスキャンによる脳函の矢状断面をみている。頭蓋天井の前頭骨と、後頭部の面(大後頭孔と基底結節を通る面)のなす角度が、シンラプトル、ギガノトサウルス、カルカロドントサウルスでは鈍角であるが、アクロカントサウルスなど多くの獣脚類ではむしろ鋭角である。これは後頭顆より下で基底後頭骨が後方に突出しているためであるという。またこれら3種では、傍後頭骨突起が強く後側方を向いていることから、後頭顆に対して顎関節の位置がより後方にあったともいっている。
このようなカルカロドントサウルス類との類似は、現在は収斂と考えられている。
(続く)
参考文献
Currie, P. J. & Zhao, X.-J. (1994). A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Canadian Journal of Earth Sciences, 30 [for 1993], 2037-2081.
Brusatte, S. L. & Sereno, P.C. (2008). Phylogeny of Allosauroidea. Journal of Systematic Palaeontology, 6, 155-182.
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300.
Coria, R. A. & Currie, P. J. (2002). The braincase of Giganotosaurus carolinii (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina. Journal of Vertebrate Paleontology, 22, 802-811.
Wikipedia (English). Metriacanthosauridae.
コメント ( 0 ) | Trackback ( 0 )
ヘピン続き

当サイトではいくつかの恐竜のイラストを描く際に、恐竜博などで実際に見た復元骨格を参考にしているが、参考にできない場合もある。このヘピンゲンシスの場合、復元骨格のポーズに難があり、描く気にならなかった。いちいち説明することもないが、胴体が水平なのに頭と首は45°上を向き、尾は垂れ下がっていた。また足の指にはパッドがあるとしても、少しつま先立ちすぎではないか等々。今回「無理やり水平化」してみたが、直しきれない点も残っている。
また、細かい話になるが、ヘピンゲンシスの復元骨格の体型(プロポーション)にも気になる点がある。2005年のジュラ紀大恐竜展では、実物化石が来ていた。頭骨と全身骨格の両方に「実物」と書かれていた。頭骨はガラスケースに入って警備員さんが見張っていた方が実物だから、全身骨格の頭はレプリカで、四肢はほとんど推定で作ったはずだが、脊椎骨や肋骨、腰帯などが実物と解釈した。脊椎骨が全部実物なら大きさが狂うはずはなく、信頼できるはずである。シンラプトルもヤンチュアノサウルスも大体アロサウルスと似た体型と思えば違和感はない。実際、この恐竜展の主役を張る堂々たる全身骨格で、胴が長めのアロサウルス体型であった。
しかし2007年の名古屋の「恐竜大陸」では、ポーズは2005年と同じだったが、ひとまわり小さくなったような気がしたが気のせいだろうか。さらに2008年の幕張の「恐竜大陸」では、ポーズも変わってゴジラ立ちというか、尾を下げて立ち上がった姿勢であった。幕張の方が会場スペースが小さかったためかもしれない。全く同じかどうかわからないが北京自然史博物館にあるキャストも立ち上がった姿勢のものである。これをみると頸が短くティラノサウルス的なプロポーションとも見える。
Gao (1992)によると、頭骨の長さは1040 mm、頸椎の合計の長さは890 mm、胴椎の合計は1550 mmとある。素直に考えると、10:9で頭骨よりも頸が短いのであり、頭が大きく、頸が短かめ、胴も短かめのティラノサウルス的な体型に思えるのだが。実際、先述のようにヤンチュアノサウルスと比べて頭が大きいのは確かで、頭骨:頸椎:胴椎が1 : 0.9 : 1.6とすると、頸椎+胴椎に対する頭骨の比率が1/2.5で40%になる。すると(実物を使っているはずの)ジュラ紀大恐竜展の復元骨格は、体が少し大きく作られているのではないか。頸と胴が少しずつ長くなっているようにも見える。それとも椎間板など関節部分の軟骨の長さを考慮すると、自然にこのようになるのだろうか。よく見ると、後方の胴椎(哺乳類の腰椎にあたる)で椎体と椎体の間に隙間が空いているが、自然にこうなるのか、意図的にしたのか。大きさの比率といっても、ポーズや見る角度によっても変わるので、本当のところは測ってみないとわからない。個々の骨は実物だが、隙間を空けるなど組み立て方によって大きくなったのか。(実物はほんの一部で、脊椎骨は10%くらい大きく作ったキャストということはないのか。)
しかし、顔に関してはジュラ紀大恐竜展の方が正確というか、実物化石に忠実な気がする。立ちポーズの方の顔は、微妙に人工的なニュアンスが感じられる。当然ながら、ジュラ紀大恐竜展の実物頭骨はすばらしかった。後眼窩骨のごつごつした表面には、トゲ状に尖った部分もみられた。名古屋の恐竜大陸ではレプリカがあったが、表面のディテールが失われていて残念であった。
このヘピンゲンシスはかなり立派な肉食恐竜だし、せっかく保存も良いのだから、願わくは正確なプロポーションで、かつ自然な姿勢の復元骨格が見たいものである。
コメント ( 3 ) | Trackback ( 0 )
シンラプトル・ヘピンゲンシス(ヤンチュアノサウルス・ヘピンゲンシス)
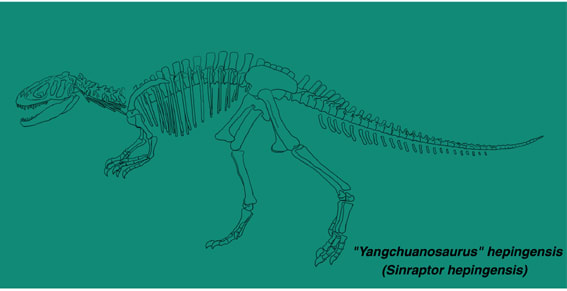
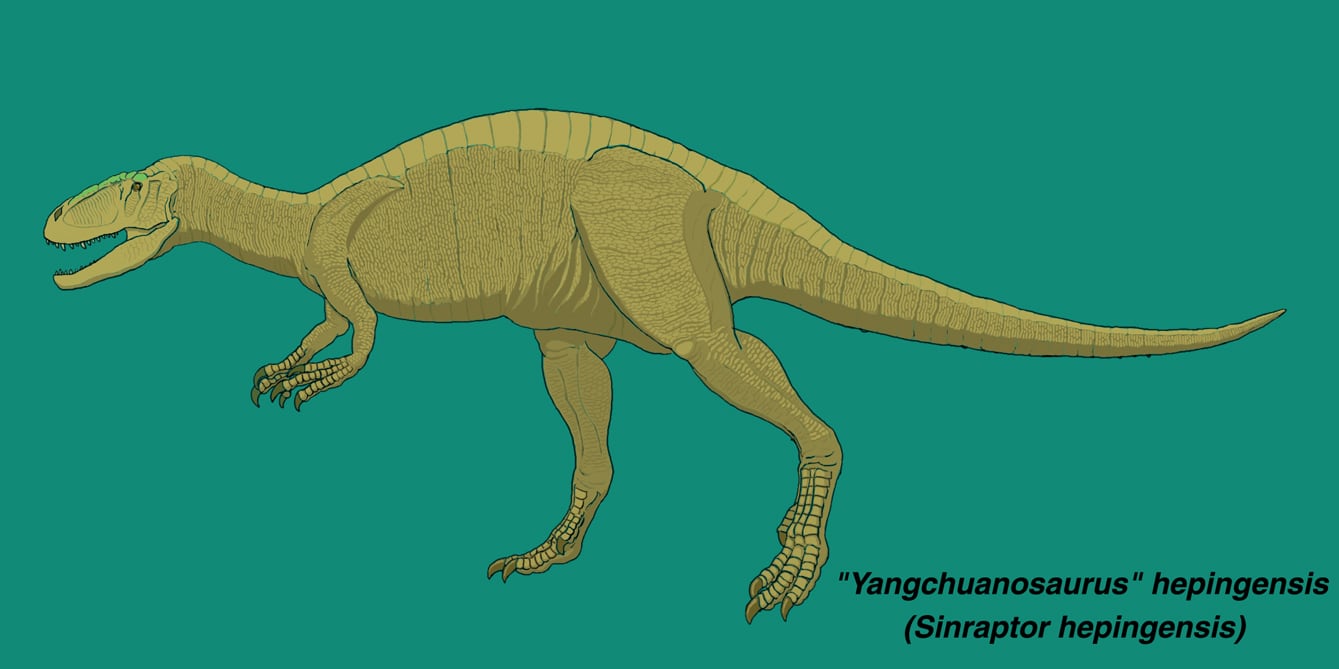
2005年の「ジュラ紀大恐竜展」で「ヤンチュアノサウルス」が来日したのは記憶に新しい。あのときの図録には、「この恐竜は現在、シンラプトルという意見もあります」という説明が付されている。しかしこれでは、最初に何をもってヤンチュアノサウルスと同定され、それがなぜ後にシンラプトルとされることになったのか、全くわからない。2007、2008年の「恐竜大陸」でも登場したが、日本ではヤンチュアノサウルスのまま紹介されている。
この恐竜は1992年に自貢恐竜博物館のGaoによってヤンチュアノサウルスの新種、Yangchuanosaurus hepingensisとして記載された。その後、1993(1994)年にCurrie and Zhaoのシンラプトル・ドンギの記載論文の中で、シンラプトル属の第2の種であろうと示唆された。ただしCurrie and Zhaoは実際の標本を観察して正式に記載したわけではないので、いまのところ"Yangchuanosaurus" hepingensisとしておくのが妥当な気もする。ところがThe Dinosauria 2nd.ed. やWikipedia(英語版)でもCurrie and Zhaoの意見を採用してすっかりSinraptor hepingensis になっている。欧米の多くの研究者にはそのように認識されていることになる。Wikipediaではフロリダ州マイアミに展示されたこの恐竜のキャストの写真があるが、Sinraptor hepingensisとあるから、アメリカではシンラプトルとして展示されたということだろうか。
自貢恐竜博物館の模式標本ZDM 0024は、完全な頭骨、9個の頸椎、14個の胴椎、5個の仙椎、35個の尾椎が関節状態で保存されている。また肩帯、腰帯もほとんど完全に保存されているが、四肢はほとんど見つかっていない。
体は大きく全長8mに達する。頭骨はがっしりしており、顔の部分は長く丈が低い。第1前眼窩窓(=前眼窩窓)は大きく前後に長く、二等辺三角形をなす。第2前眼窩窓(=Maxillary fenestra)は小さく四角形である。その他に上顎骨にはMaxillary depressionという窪みがあり、これは長い楕円形である。頭頂骨の後方突起がよく発達している。上後頭骨は幅が狭く、正中の稜がよく発達している。涙骨は傾斜している。歯骨は厚く頑丈で、比較的丈が高い。歯は比較的小さく、前上顎骨と上顎骨の歯は薄い。前上顎骨歯は4本、上顎骨歯は13本、歯骨歯は16本ある。
9個の頸椎は後凹型で、後方の頸椎には腹側の稜がある。14個の胴椎は、比較的短い両扁平型の椎体と、丈の高い板状の神経棘をもつ。5個の仙椎は互いに癒合している。尾椎は両凹型で、中央と後方の尾椎には長い前関節突起がある。腸骨は丈が高く前方突起が腹側に曲がっている。恥骨のpubic footは短く幅広い。座骨の遠位端は拡がっている。
Gao (1992)によると、この標本は以下のような多くのヤンチュアノサウルスの特徴を示すという。頭骨は頑丈で、長さと高さの比率が中程度であり、6対の開口部をもつ。外鼻孔は卵形である。下側頭窓は中くらいの大きさで、その最も前後に長い部分は腹側中央にある(下ぶくれの形)。上顎骨には窪み(Maxillary depression)がある。頭頂骨にはよく発達した後方突起がある。前頭骨と頭頂骨が癒合している。下顎窓は大きい。前上顎骨歯は4本で、上顎骨歯は15本を超えない。頸椎は後凹型で、胴椎は両扁平型である。胴椎の神経棘は長方形で丈が高い。5個の仙椎は互いに癒合している。尾椎は両凹型で、前方の尾椎の神経棘は丈が高い。腰帯は頑丈で、pubic footは中程度に発達している。これらの特徴から、ヤンチュアノサウルスに属するとしている。
しかし、この標本にはそれまでに発見されていたヤンチュアノサウルスとは異なる点も多くみられた。例えば、この“ヤンチュアノサウルス”・ヘピンゲンシスとヤンチュアノサウルス・シャンヨウエンシスが異なる点は、以下のようである。仙前椎(頸椎+胴椎)の長さに対する頭骨の長さの比率が、ヘピンゲンシスでは41%であるが、シャンヨウエンシスでは31%である。第1前眼窩窓は、ヘピンゲンシスでは前後に長い二等辺三角形であるが、シャンヨウエンシスでは正三角形である。第2前眼窩窓は、ヘピンゲンシスでは長方形であるが、シャンヨウエンシスでは三角形である。歯はヘピンゲンシスでは比較的小さく扁平であるが、シャンヨウエンシスでは比較的大きく太い。歯骨はヘピンゲンシスでは比較的丈が高いが、シャンヨウエンシスでは丈が低い。胴椎はヘピンゲンシスでは14個で椎体が短いが、シャンヨウエンシスでは13個で椎体が比較的長い、などである。またヤンチュアノサウルス・マグヌスとも比較している。これらの特徴から、ヤンチュアノサウルス属の新種、ヤンチュアノサウルス・ヘピンゲンシスとされた。
Currie and Zhao (1993)は3種のヤンチュアノサウルスのうち、ヘピンゲンシスをシンラプトルに属するとした。シンラプトルはヤンチュアノサウルスと比較して頭骨がより長く、丈が低いこと、下側頭窓が大きいこと、後眼窩骨の側頭窓間突起が鱗状骨に覆われていること、後眼窩骨の粗面が発達して涙骨と後眼窩骨の間がほとんど閉じていること、などで区別できるとしている。
参考文献
Gao, Y. (1992) Yangchuanosaurus hepingensis - a new species of carnosaur from Zigong, Sichuan. Vertebrata PalAsiatica 30: 313-324.
(Translation of Gao (1992) was by J. Jin and obtained courtesy of the Polyglot Paleontologist website (http://www.paleoglot.org).)
Currie, P. J., and X.-J. Zhao. (1993) A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, Peopleユs Republic of China. Canadian Journal of Earth Sciences 30: 2037-2081.
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ |