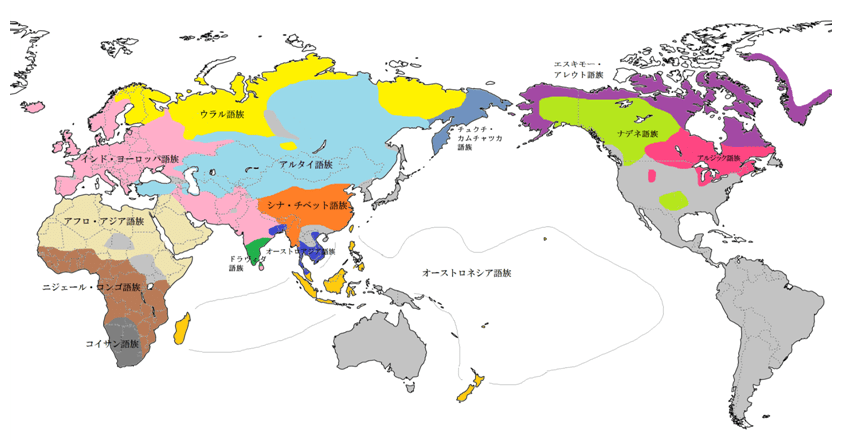
次に、言語の分布をみてみよう。世界の言語は様々な分布様相を示す。インド・ヨーロッパ語族やアルタイ語族のようにユーラシア大陸を広く覆う言語群が存在する一方、北海道のアイヌ語やスペイン・フランス国境のバスク語というような系統不明な言語が孤立して残存していたり、ニューギニア島やアメリカ先住民の言語のように互いの系統が不詳な言語が多く共存している地帯も存在する。世界の言語の分布様式は一般に1)モザイク分布 2)均質分布 3)残存分布 の概ね3型に区別できると思われる。
1)
モザイク分布-は互いに系統不詳あるいは証明が困難な言語が狭い分布で多数共存する状態である。南米のアメリカ先住民の言語やニューギニア島の言語がそうである。これらの地域について一般に言えることは狩猟採集生活を行い、人類入植以来ある程度の期間がたっていながら、一度も言語的に単一化された歴史がない、広範囲の支配>被支配の関係が生じなかった地域ということができる。もちろん「国」というものが存在せず、言語的にも互いに独自性を強めてきたのである。
2)
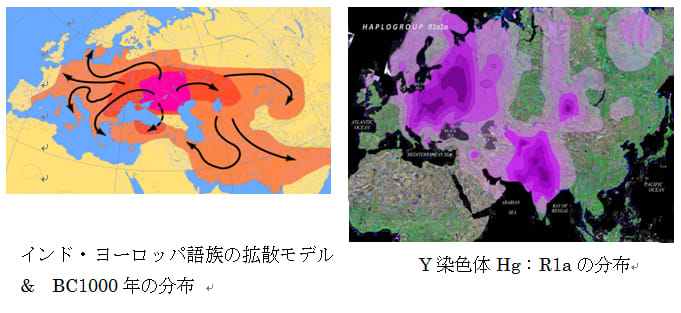
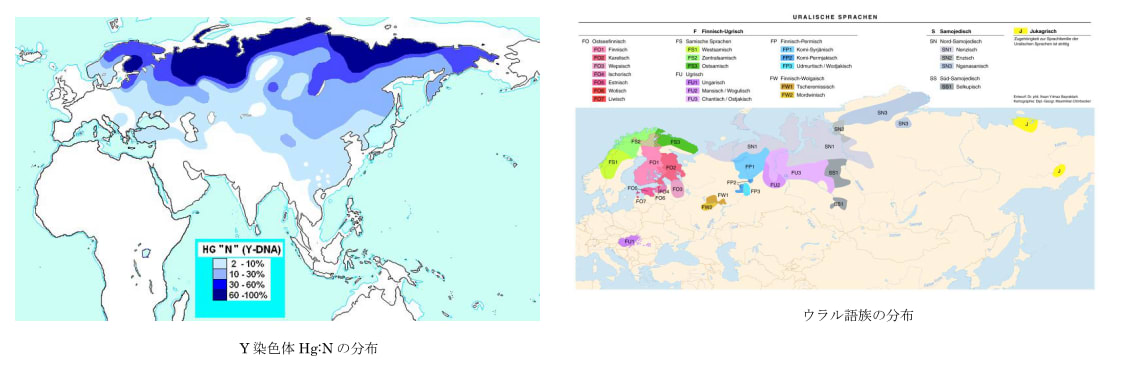
均質分布-はモザイク分布と対極的で、ごく最近の単一語族の急速拡散の結果である。ある集団が文化的優越性を生じる要素(農耕、騎馬など)を身につけた場合、周辺の狩猟採集民などを駆逐、同化して言語を置き換え、分布を広げることで急速拡散をする場合である。1万年ほど前に農耕が開始して以降農耕民の急速拡散が生じた。古メソポタミア語族、古西欧語族(共に畑作)、オーストロアジア語族(稲作)などが分布を広げたと考えられる。6000年前頃になると騎馬戦士文化を伴ったインド・ヨーロッパ語族が拡散し、5000~4000前頃からはアルタイ語族(騎馬文化)やウラル語族(トナカイ遊牧)などが分布を広げたようである。また優れた航海術を身につけ台湾からインドネシア、太平洋地域に広く拡散したオーストロネシア語族もこれに含まれる。
3)
残存分布-はかつては均質分布であった言語が、新たに拡大してきた均質分布の言語に置き換えられ、駆逐、分断されるかたちの言語である。かつてパキスタン~インドに広く分布し、インド・ヨーロッパ語族の進入で分断したドラヴィダ語族や、かつて東南アジアに広く分布し現在ではタイ系言語やビルマ系言語に分断される形となっているオーストロアジア語族などがあげられる。この分布は時間軸による語族の栄枯盛衰を反映していると言える。
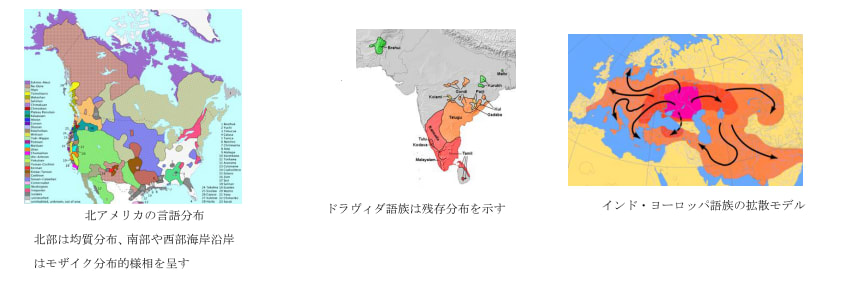
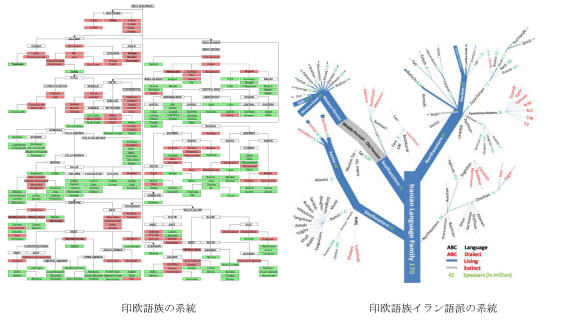
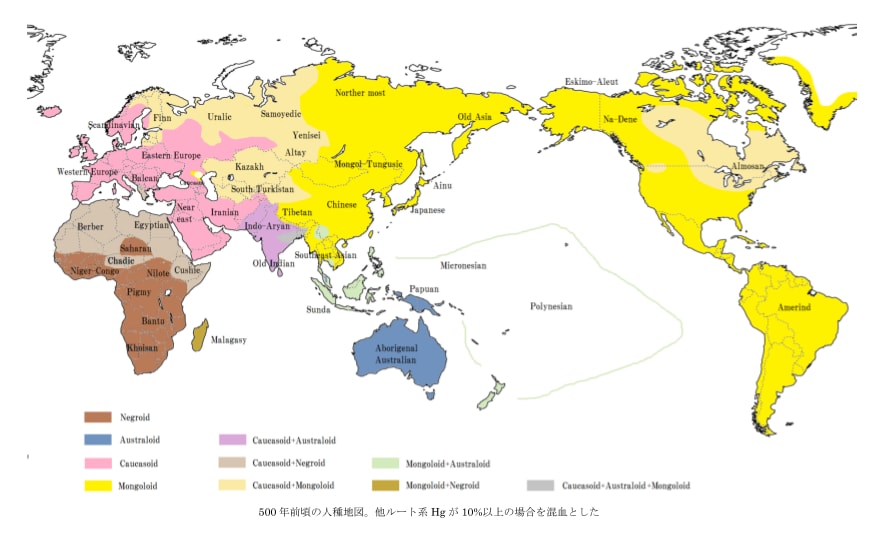
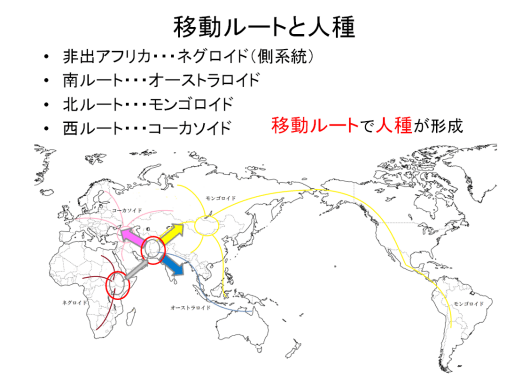
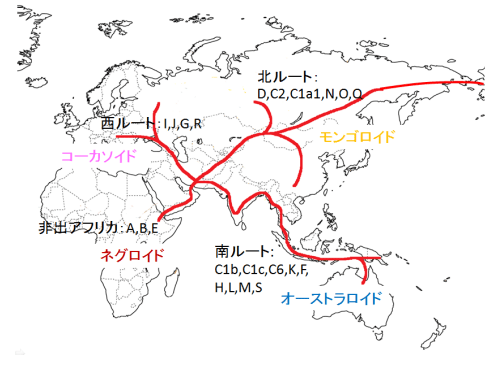
図19、20、21(Wipikedia)
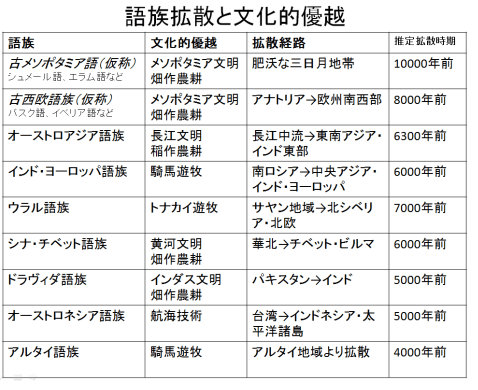
世界の語族は歴史学的、考古学的におよそ拡散経路、時期が推定されているが、一欄を表1-1に示した。語族拡散の必要なのは文化的優越であり、農耕、騎馬戦闘、航海術などが考えられる。約1万年前に開始された農耕を携えて拡散していった語族がいくつか見られ、次段階では騎馬遊牧などより進んだ文化的優越性を持つ語族が拡散していったと考えられる。
表1-1
ところで、高橋ほか(1995)によると文化地理学では伝播を大きくa1)接触拡大伝播 a2)階層性拡大伝播 b)移転伝播の3種に大別できるという。a1)接触拡大伝播は紙に水が浸み込んでいくように接触によってじわじわと拡大するものである。方言語彙の「方言周圏論」が例としてあげられる。a2)階層性拡大伝播は重要人物が他の重要人物へ、大都市から大都市へというように、他の人々を飛び越えて伝播する過程である。これは世界各地における近代の西欧文化の伝播などが当てはまるであろう。b)移転伝播は文化特性を持つ個人や集団が新しい居住地へ移動することで発生する。ヨーロッパ人がアメリカ大陸に移住することによる完全な西欧化はまさに好例であろう。
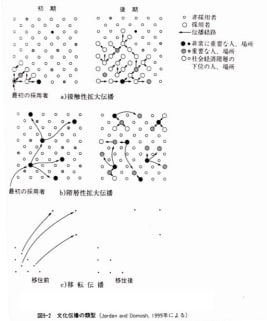
図1-22
さて、これらを言語と住民という視点に応用してみよう。言語の伝播、拡散形式にはさまざまなものが考えられる。外部から強力な支配者の支配を受ければ、住民がほとんど変わらず言語だけが支配者のものに置き換わることもありうるが、一方で言語を使用するのは人間である以上、人間の混合が起こらない場合、語彙の借用などを除き言語は本質的には変化しないようにも思える。言語と住民の関係には場所を基準にした場合、大きく以下の5型に分類できると思われる。
1.
言語交換(住民そのまま)(Dominant minority)
住民は殆ど変化せず外部からの少数の支配者層の母語に言語交換する場合。
例)トルコ共和国:トルコ語…アナトリア地方は古来より数回の言語交換を受けた
2.
住民混合+言語交換
外部からの支配者層と原住民が混合した状態であり、言語は支配者層のものに交換した場合。
例)インド:インド・アーリア語
3.
住民混合+言語混合
外部から進入した集団と原住民が混合し、言語も混合した場合。
例)ハザーラ語(モンゴル語+ペルシャ語)。日本語(族)もこのタイプであると思われる。
4.
住民交換+言語交換
外部から進入した集団が原住民をほとんど駆逐してしまう場合。
例)北海道:アイヌ⇒日本人…北海道はアイヌの土地であったが、現在は99%以上日本人が居住している。
文化地理学の伝播拡散モデルに当てはめると、1が階層性拡大伝播 2~4が移転伝播に近いと言える。なおこれらは質的概念であり絶対的基準はない。
→次頁「人種・民族・言語・遺伝子」へ
<文献>
高橋伸夫ほか(1995)『文化地理学入門』東洋書林