東アジアの人類集団についてまずおさえておくべき前提として、集団の流れとしては専ら北から南の方向であるということがある。前に紹介したYali Xueら(2006)によるY染色体の多様性による集団の拡散年代解析(P9図1-4)から 、北部集団の拡散年代が古く、南部が新しい。また、ミトコンドリアDNAハプログループについて、北部はD*,D4*,C*など未分化パラグループが多いのに対し、南部ではR4a1a,R9c,B4a2,F1a3,F1a4,F3,M7b3,N9a6,Y2,E など特定の下位グループのみが展開する傾向がみられる(崎谷2009)。これらより、東アジアの人類集団の流れは専ら北から南への方向であったと結論付けることができる。出アフリカ後に北ルートをとったモンゴロイド人種のホームランドはモンゴル高原、アルタイ山脈付近であり、その一部が比較的最近になって南下していったものと考えられる。東アジア北部が寒冷にもかかわらず人口集積地だったのは、数万年前は草原が広がり食料となる大型哺乳類が豊富だったからであろう。

図3-1
では、東アジアの植生について見てみよう。気候学では北方ほど低温、内陸部ほど乾燥というのが原則である。中緯度では高圧帯により砂漠ができやすいが、大陸東岸は湿潤であり、東アジアの中緯度海岸部に砂漠は存在しない。東アジア海岸部の植生は南から熱帯多雨林、照葉樹林、夏緑樹林、針葉樹林となる。北緯45度付近の内陸部では降水量が減少しステップや砂漠が広がる。また標高4000mのチベット高原は高山植生となっている。
東アジアの生業も概ね植生に対応している。最も顕著なのはステップ・砂漠地帯で、専ら遊牧生活が営まれ、彼らは騎馬民族であり、アルタイ語族に属す。チベット高原地帯ではヤクの放牧を中心とした牧畜が営まれている。北部の針葉樹林では多くが狩猟採集生活を行い、北緯55度以北になるとトナカイ遊牧を行う民族が大勢を占めるようになる。中国北部の夏緑樹林地域は畑作地帯で、古くから麦や雑穀を栽培してきた。そして、中国中部~東南アジア、日本など広大な分布域を持つ稲作農耕がある。稲は人口扶養力が大きく、東アジア地域を特徴づける生業といっていいであろう。
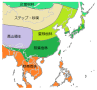 図3-2
図3-2 図3-3
図3-3
このような環境、生業を背景に、東アジアでは長江文明、黄河文明、遼河文明という3文明が興った。遼河文明および黄河文明は冷涼、乾燥気候下で麦・雑穀作を基盤とし、ブタなどの牧畜も行う。対して長江文明はアジアモンスーン多雨気候の元で専ら稲作に依り、漁撈を併存する(安田2009)。起源は長江文明が古く、最古の稲作遺跡は中国江西省で約12000前のものが発見されている。
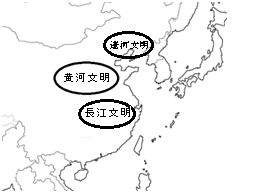
図3-4
黄河文明の構成民族はシナ・チベット語族(Y染色体Hg:O3)と考えられる。華北が発祥であるが、チベット・ビルマ語派はその後南進し、ビルマやチベットに分布を広げたと考えられる。人口13億を誇る漢民族も黄河文明から興った。
一方の長江文明は複数系統の民族が構成員であり、オーストロアジア語族(O2a)、タイ・カダイ語族、オーストロネシア祖族(O1など)、倭人(O2b)が担い手であったと考えられる。
そして遼河文明の担い手はウラル語族(N1*,N1c)と考えられる(Yuli et al.2013)。彼らはシベリアから北欧にかけて広く拡散したが、その祖地である遼河地域では気候変動()および後の漢民族の膨張とアルタイ系騎馬民族の侵攻に追われることとなった。
漢民族は黄河中流域の「中原」を発祥とする民族であるが、有史以来拡大膨張してきた。特に気候が寒冷化した2500年前頃(春秋時代)に大規模な南下を起こす。この時代、中国には漢民族の国家のほかに、呉、越など別民族の国家も存在した。呉、越は長江文明を担った人々と考えられるが、漢民族の膨張南下により2500年前頃に長江文明は崩壊し、呉の民の一部は日本へ逃れ倭人となったようである。(崎谷2008 澤田1999)。漢民族の南下に伴う異民族の混合・駆逐は長江文明崩壊以降も断続的に行われてきたようである(崎谷2009)。現在も中国南部に少数民族が居住するが、かつては中南部の広範囲にわたり非漢民族の人々が多数居住していたと考えられる。
長江文明の崩壊以前から東南アジアでは現在の中国地域から断続的な民族移動が行われてきた。農耕開始以前は東南アジアには先住民オーストラロイド(南ルート系)とわずかなモンゴロイド(北ルート系)が共存していたと思われるが、稲作開始に伴い長江流域から民族移動が始まった。
最初に東南アジアに移住してきたのはオーストロアジア語族(O2a)である。インド東部~ベトナムにかけて分断分布を示すこの語族は稲作を携えて東南アジアに拡散した最古層の住民と思われ、インドに稲作を伝達したのも彼らと考えられる。拡散年代は6300年前頃との説があり、(Ilia Peiros 2004)、インド最古の稲作遺跡は5000年前頃のものが発見されている。
2番目に移動を開始したのはオーストロネシア語族であり、華南から台湾へ6000年前頃に移住したのち、台湾からフィリピン、インドネシア方面へ拡散していった。およそ5000年前と考えられている(Kun, Ho Chuan 2006 Bellwood 1989)。
3番目の移住者はチベット・ビルマ系民族で、かつてはその一派は羌と呼ばれた。四川・雲南地域からビルマ回廊を南下していき、ビルマ平野部では雑穀農耕から稲作農耕へと生業を切り替えた(池橋2005)。おおよそ4000年前頃と考えられる(崎谷2009)。
最後に移住してきたのはタイ・カダイ語族である。タイ系民族はおそらく春秋時代の百越と呼ばれた人々の一部で、長江文明崩壊に伴い長江流域から現在のタイまで南下してきたと考えられる(池橋2005)。2500年前以降のことであり、東南アジアで最も新しい民族ということになる。
もっとも、近年は中国人の東南アジア流入もあり、この限りではないが、有史以来漢民族の東南アジア流入は多かれ少なかれ存在してきたと思われる。このように東アジアでは常に北から南への移動が繰り返されてきた。

図3-5
表3-1

最後に、東アジアの歴史をみていく上で欠かすことのできない東西交流について述べておきたい。古来より東西アジアの交流は少なからず存在した。中国北部の麦作は古代メソポタミアから伝達したものであろう。有史以降とりわけ東西交流を担ってきたのは騎馬民族である。ユーラシア内陸ステップ地帯を疾走する騎馬民族は並はずれた機動力により東西交流の媒体となってきた。古くはインド・ヨーロッパ語系クルガン、スキタイに始まり、アルタイ語族系チュルク、モンゴル族が騎馬文化を受け継いた。チンギスハーンが築いたモンゴル帝国やヨーロッパを震撼させたフン族など、ユーラシア大陸において騎馬民族がいかに大きな影響を持ったかがわかる。万里の長城は中国の歴代王朝が騎馬民族南下を防ぐために造ってきたものである。
現在のトルコ共和国にかつて存在したヒッタイト(インド・ヨーロッパ語族)は当時世界一の製鉄技術を持つ集団であったが、騎馬民族が媒体となり東アジアにも製鉄が伝達された。日本においては古代出雲王朝が製鉄術に長けていたとされるが、草原の道を通って騎馬民族により伝達されたものであろう。
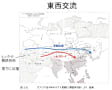
図3-6
<文献>

図3-1
では、東アジアの植生について見てみよう。気候学では北方ほど低温、内陸部ほど乾燥というのが原則である。中緯度では高圧帯により砂漠ができやすいが、大陸東岸は湿潤であり、東アジアの中緯度海岸部に砂漠は存在しない。東アジア海岸部の植生は南から熱帯多雨林、照葉樹林、夏緑樹林、針葉樹林となる。北緯45度付近の内陸部では降水量が減少しステップや砂漠が広がる。また標高4000mのチベット高原は高山植生となっている。
東アジアの生業も概ね植生に対応している。最も顕著なのはステップ・砂漠地帯で、専ら遊牧生活が営まれ、彼らは騎馬民族であり、アルタイ語族に属す。チベット高原地帯ではヤクの放牧を中心とした牧畜が営まれている。北部の針葉樹林では多くが狩猟採集生活を行い、北緯55度以北になるとトナカイ遊牧を行う民族が大勢を占めるようになる。中国北部の夏緑樹林地域は畑作地帯で、古くから麦や雑穀を栽培してきた。そして、中国中部~東南アジア、日本など広大な分布域を持つ稲作農耕がある。稲は人口扶養力が大きく、東アジア地域を特徴づける生業といっていいであろう。
 図3-2
図3-2 図3-3
図3-3このような環境、生業を背景に、東アジアでは長江文明、黄河文明、遼河文明という3文明が興った。遼河文明および黄河文明は冷涼、乾燥気候下で麦・雑穀作を基盤とし、ブタなどの牧畜も行う。対して長江文明はアジアモンスーン多雨気候の元で専ら稲作に依り、漁撈を併存する(安田2009)。起源は長江文明が古く、最古の稲作遺跡は中国江西省で約12000前のものが発見されている。
図3-4
黄河文明の構成民族はシナ・チベット語族(Y染色体Hg:O3)と考えられる。華北が発祥であるが、チベット・ビルマ語派はその後南進し、ビルマやチベットに分布を広げたと考えられる。人口13億を誇る漢民族も黄河文明から興った。
一方の長江文明は複数系統の民族が構成員であり、オーストロアジア語族(O2a)、タイ・カダイ語族、オーストロネシア祖族(O1など)、倭人(O2b)が担い手であったと考えられる。
そして遼河文明の担い手はウラル語族(N1*,N1c)と考えられる(Yuli et al.2013)。彼らはシベリアから北欧にかけて広く拡散したが、その祖地である遼河地域では気候変動()および後の漢民族の膨張とアルタイ系騎馬民族の侵攻に追われることとなった。
漢民族は黄河中流域の「中原」を発祥とする民族であるが、有史以来拡大膨張してきた。特に気候が寒冷化した2500年前頃(春秋時代)に大規模な南下を起こす。この時代、中国には漢民族の国家のほかに、呉、越など別民族の国家も存在した。呉、越は長江文明を担った人々と考えられるが、漢民族の膨張南下により2500年前頃に長江文明は崩壊し、呉の民の一部は日本へ逃れ倭人となったようである。(崎谷2008 澤田1999)。漢民族の南下に伴う異民族の混合・駆逐は長江文明崩壊以降も断続的に行われてきたようである(崎谷2009)。現在も中国南部に少数民族が居住するが、かつては中南部の広範囲にわたり非漢民族の人々が多数居住していたと考えられる。
長江文明の崩壊以前から東南アジアでは現在の中国地域から断続的な民族移動が行われてきた。農耕開始以前は東南アジアには先住民オーストラロイド(南ルート系)とわずかなモンゴロイド(北ルート系)が共存していたと思われるが、稲作開始に伴い長江流域から民族移動が始まった。
最初に東南アジアに移住してきたのはオーストロアジア語族(O2a)である。インド東部~ベトナムにかけて分断分布を示すこの語族は稲作を携えて東南アジアに拡散した最古層の住民と思われ、インドに稲作を伝達したのも彼らと考えられる。拡散年代は6300年前頃との説があり、(Ilia Peiros 2004)、インド最古の稲作遺跡は5000年前頃のものが発見されている。
2番目に移動を開始したのはオーストロネシア語族であり、華南から台湾へ6000年前頃に移住したのち、台湾からフィリピン、インドネシア方面へ拡散していった。およそ5000年前と考えられている(Kun, Ho Chuan 2006 Bellwood 1989)。
3番目の移住者はチベット・ビルマ系民族で、かつてはその一派は羌と呼ばれた。四川・雲南地域からビルマ回廊を南下していき、ビルマ平野部では雑穀農耕から稲作農耕へと生業を切り替えた(池橋2005)。おおよそ4000年前頃と考えられる(崎谷2009)。
最後に移住してきたのはタイ・カダイ語族である。タイ系民族はおそらく春秋時代の百越と呼ばれた人々の一部で、長江文明崩壊に伴い長江流域から現在のタイまで南下してきたと考えられる(池橋2005)。2500年前以降のことであり、東南アジアで最も新しい民族ということになる。
もっとも、近年は中国人の東南アジア流入もあり、この限りではないが、有史以来漢民族の東南アジア流入は多かれ少なかれ存在してきたと思われる。このように東アジアでは常に北から南への移動が繰り返されてきた。
図3-5
表3-1
最後に、東アジアの歴史をみていく上で欠かすことのできない東西交流について述べておきたい。古来より東西アジアの交流は少なからず存在した。中国北部の麦作は古代メソポタミアから伝達したものであろう。有史以降とりわけ東西交流を担ってきたのは騎馬民族である。ユーラシア内陸ステップ地帯を疾走する騎馬民族は並はずれた機動力により東西交流の媒体となってきた。古くはインド・ヨーロッパ語系クルガン、スキタイに始まり、アルタイ語族系チュルク、モンゴル族が騎馬文化を受け継いた。チンギスハーンが築いたモンゴル帝国やヨーロッパを震撼させたフン族など、ユーラシア大陸において騎馬民族がいかに大きな影響を持ったかがわかる。万里の長城は中国の歴代王朝が騎馬民族南下を防ぐために造ってきたものである。
現在のトルコ共和国にかつて存在したヒッタイト(インド・ヨーロッパ語族)は当時世界一の製鉄技術を持つ集団であったが、騎馬民族が媒体となり東アジアにも製鉄が伝達された。日本においては古代出雲王朝が製鉄術に長けていたとされるが、草原の道を通って騎馬民族により伝達されたものであろう。

図3-6
<文献>
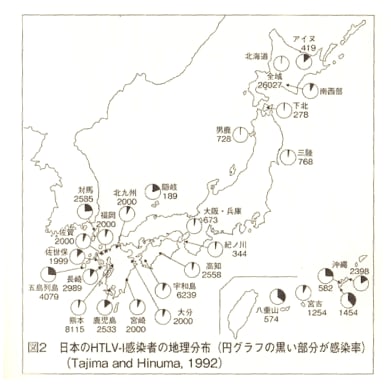
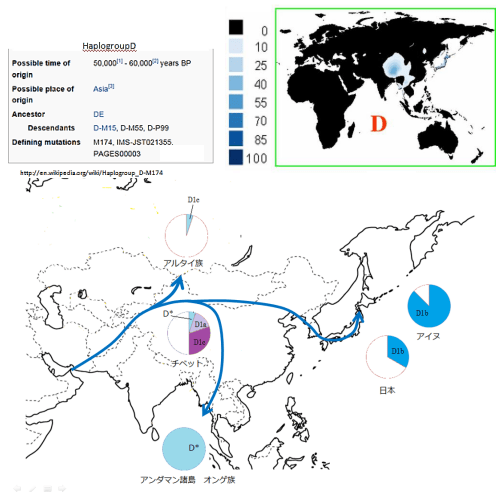
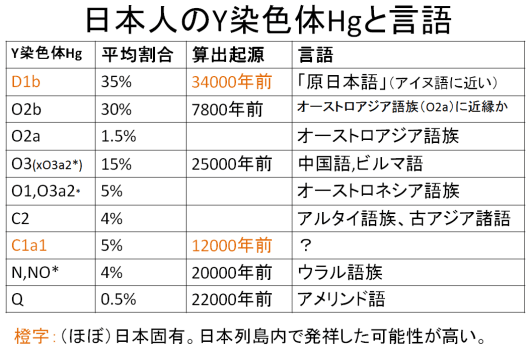
 図2-33、34
図2-33、34