








HaplogroupD1b(D-M55)
ハプログループDは日本列島、チベット、アンダマン諸島などで高頻度に観察されている。非出アフリカ系統のハプログループEと姉妹関係を成し、起源は55000年前程と非常に古い。起源年代とその分布様相から、かつては東アジアを広く覆っていたが、駆逐され島国日本と高山チベットに取り残されたようである。ハプログループDはモンゴロイドを形成する集団の最古層に位置し、イラン→アルタイ山脈→東アジアという北ルートで東アジアに達したと考えられる(崎谷2009)。
日本列島に分布するのはHg:D1b(D-M55)で、他に朝鮮で0~2%程確認されるのみでほとんど日本列島固有である。D1bは日本人に25~45%,琉球人に55%,アイヌに85%の頻度で確認されている。D1bの起源年代は33700±2200年前(She et al. 2003)で、日本列島に人類が居住し始めた年代とほぼ一致する。
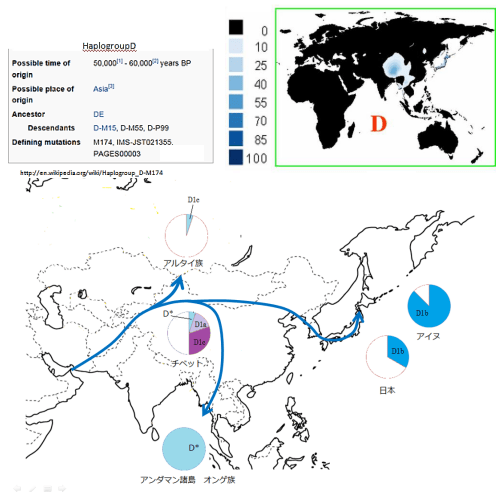
図2-6、図2-7、表2-5

図2-8
HaplogroupO1(O-MSY2.2)
このハプログループはオーストロネシア語族と関係し台湾先住民で66%を占める。東南アジアのタイ・カダイ語族にも比較的高頻度で確認される。ただし両語族ともハプログループO3も共担する。日本にはO1が0%~4.2%(Nonaka 2007)ほど確認されている。
HaplogroupO2a(O-PK4),O2b(O-M176)
Hg:O2aは東南アジアで高頻度にみられ、オーストロアジア語族と関連している。日本では2.4%観察されたという報告がある(Hammer2006)。
Hg:O2bは日本、朝鮮、満州に高頻度で観察され、ベトナムやタイでも確認される。O2bの起源は7800年前と推定される(Katoh 2004)。O2bは日本で平均30%程観察され、D1bと共に日本人の2大Hgとなっている。
共に長江文明の担い手である。

図2-9、図2-10、図2-11、表2-5
HaplogroupO3(O-M122)
東アジアに広く分布し、下位グループも多い。北部漢民族では60%以上の高頻度で観察される。O3a2c1がシナ・チベット語族を担い、特にO3a2c1aがチベット・ビルマ語派を担っている。O3a2*などいくつかのブランチはオーストロネシア語族と関連しているようである。O3a2c1と関連するシナ・チベット語族は雑穀・麦作農耕の黄河文明の担い手である。日本にはO3全体で20%程が確認されるが、下位グループの分析が追い付いていないようである。

図2-12、図2-13
HaplogroupC1a1(C-M8)
このハプログループの最大の特徴は日本固有であることである。日本全国で平均5%ほど確認されるが、国外では一切観察されていない。つまり日本列島内で祖形C1a*から発祥した可能性が高いと思われる。遺伝学者の計算で発祥年代が12000年前後(Hammer et al.2006)とされており、縄文文化の開化とリンクしていることが考えられる。
表2-6

HaplogroupC2(C-M217)
このハプログループはユーラシア中部~北米に多い。アルタイ語族、古アジア諸語、ナデネ語族と関連している。日本には平均4%ほど観察され、日本固有のC2aも存在する。福岡で7%以上と高い(Hammer et al.2006)。アイヌでも12%観察されている。起源地はアルタイ・サヤン地域と推定される(Zegura et al.2004)。

図2-14、表2-7

図2-15
HaplogroupN (N-M231)
このハプログループはユーラシア北部(極北地域)に多い。分布域は北東シベリアから北欧にまで及ぶ。日本では平均4%ほど観察され、特に青森で7%以上と高い(Hammer et al.2006)。N1cはウラル語族と関連している。起源地は中国北東部と考えられる。遼河文明の担い手である(Yuli et al.2013)。

図2-16、表2-8
HaplogroupQ(Q-M242)
このハプログループはアメリカ先住民の大半を占める。ユーラシアにおいてはケット族に90%以上、セルクプ族に60%以上認められるが総じて低頻度である。イラン付近で発祥し急速にシベリアを移動しアメリカへ入ったようである。日本人には約0.5%観察される。

図2-17
日本人の Y 染色体の起源年代と言語の関係
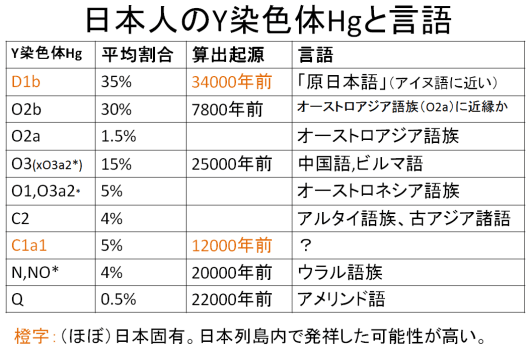
以上紹介した日本人の Y 染色体の頻度、起源年代、関係言語をまとめたものが下表である。最も多い Hg:D1b は起源 34000 年前と最も古く、しかもほとんど日本列島固有であるから、列島内発祥の可能性も高く、日本人の最古層と考えられる。これが主に縄文時代人の中核を占めていたようである。彼らの言語を「原日本語」と呼びたい。大陸部でほとんど駆逐されてしまった Hg:D の系統が高頻度で残存していることは、日本における縄文から弥生への移行には武力征服はほとんどなく、平和のうちに推移したと推定できる。ハプロルグープ O の下位群とはおもに弥生時代以降に流入したようであるが、以下、考古学的、歴史的側面からもこの考えを検証していく。
表2-9
→次頁「日本人のミトコンドリアDNAハプログループ」
〈文献〉
・Hammer et al. 2006. Dual origins of the Japanese: common ground for
hunter-gatherer and farmer Y chromosomes.
J Hum Genet. 2006;51(1):47-58.
Epub 2005 Nov 18.
・Shi,Hong , Hua Zhong, Yi Peng, Yong-Li Dong, Xue-Bin Qi, Feng Zhang, Lu-Fang Liu, Si-Jie Tan, Runlin Z Ma, Chun-Jie Xiao, R Spencer Wells, Li Jin, and Bing Su. 2008. Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations. BMC Biol. 2008; 6: 45. Published online 2008 October 29. doi: 10.1186/1741-7007-6-45
・Nonaka,I et al 2007, Y-chromosomal Binary Haplogroups in the Japanese Population and their Relationship to 16 Y-STR Polymorphisms
・Katoh,Toru 2004, Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis
・Yali Xue et al. 2013. Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times. Genetics.December 2013, 195 (4)
・Zegura SL1, Karafet TM, Zhivotovsky LA, Hammer MF.High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas.Mol Biol Evol. 2004 Jan;21(1):164-75. Epub 2003 Oct 31.
ハプログループDは日本列島、チベット、アンダマン諸島などで高頻度に観察されている。非出アフリカ系統のハプログループEと姉妹関係を成し、起源は55000年前程と非常に古い。起源年代とその分布様相から、かつては東アジアを広く覆っていたが、駆逐され島国日本と高山チベットに取り残されたようである。ハプログループDはモンゴロイドを形成する集団の最古層に位置し、イラン→アルタイ山脈→東アジアという北ルートで東アジアに達したと考えられる(崎谷2009)。
日本列島に分布するのはHg:D1b(D-M55)で、他に朝鮮で0~2%程確認されるのみでほとんど日本列島固有である。D1bは日本人に25~45%,琉球人に55%,アイヌに85%の頻度で確認されている。D1bの起源年代は33700±2200年前(She et al. 2003)で、日本列島に人類が居住し始めた年代とほぼ一致する。
図2-6、図2-7、表2-5

図2-8
HaplogroupO1(O-MSY2.2)
このハプログループはオーストロネシア語族と関係し台湾先住民で66%を占める。東南アジアのタイ・カダイ語族にも比較的高頻度で確認される。ただし両語族ともハプログループO3も共担する。日本にはO1が0%~4.2%(Nonaka 2007)ほど確認されている。
HaplogroupO2a(O-PK4),O2b(O-M176)
Hg:O2aは東南アジアで高頻度にみられ、オーストロアジア語族と関連している。日本では2.4%観察されたという報告がある(Hammer2006)。
Hg:O2bは日本、朝鮮、満州に高頻度で観察され、ベトナムやタイでも確認される。O2bの起源は7800年前と推定される(Katoh 2004)。O2bは日本で平均30%程観察され、D1bと共に日本人の2大Hgとなっている。
共に長江文明の担い手である。
図2-9、図2-10、図2-11、表2-5
HaplogroupO3(O-M122)
東アジアに広く分布し、下位グループも多い。北部漢民族では60%以上の高頻度で観察される。O3a2c1がシナ・チベット語族を担い、特にO3a2c1aがチベット・ビルマ語派を担っている。O3a2*などいくつかのブランチはオーストロネシア語族と関連しているようである。O3a2c1と関連するシナ・チベット語族は雑穀・麦作農耕の黄河文明の担い手である。日本にはO3全体で20%程が確認されるが、下位グループの分析が追い付いていないようである。
図2-12、図2-13
HaplogroupC1a1(C-M8)
このハプログループの最大の特徴は日本固有であることである。日本全国で平均5%ほど確認されるが、国外では一切観察されていない。つまり日本列島内で祖形C1a*から発祥した可能性が高いと思われる。遺伝学者の計算で発祥年代が12000年前後(Hammer et al.2006)とされており、縄文文化の開化とリンクしていることが考えられる。
表2-6

HaplogroupC2(C-M217)
このハプログループはユーラシア中部~北米に多い。アルタイ語族、古アジア諸語、ナデネ語族と関連している。日本には平均4%ほど観察され、日本固有のC2aも存在する。福岡で7%以上と高い(Hammer et al.2006)。アイヌでも12%観察されている。起源地はアルタイ・サヤン地域と推定される(Zegura et al.2004)。
図2-14、表2-7
図2-15
HaplogroupN (N-M231)
このハプログループはユーラシア北部(極北地域)に多い。分布域は北東シベリアから北欧にまで及ぶ。日本では平均4%ほど観察され、特に青森で7%以上と高い(Hammer et al.2006)。N1cはウラル語族と関連している。起源地は中国北東部と考えられる。遼河文明の担い手である(Yuli et al.2013)。
図2-16、表2-8
HaplogroupQ(Q-M242)
このハプログループはアメリカ先住民の大半を占める。ユーラシアにおいてはケット族に90%以上、セルクプ族に60%以上認められるが総じて低頻度である。イラン付近で発祥し急速にシベリアを移動しアメリカへ入ったようである。日本人には約0.5%観察される。
図2-17
日本人の Y 染色体の起源年代と言語の関係
以上紹介した日本人の Y 染色体の頻度、起源年代、関係言語をまとめたものが下表である。最も多い Hg:D1b は起源 34000 年前と最も古く、しかもほとんど日本列島固有であるから、列島内発祥の可能性も高く、日本人の最古層と考えられる。これが主に縄文時代人の中核を占めていたようである。彼らの言語を「原日本語」と呼びたい。大陸部でほとんど駆逐されてしまった Hg:D の系統が高頻度で残存していることは、日本における縄文から弥生への移行には武力征服はほとんどなく、平和のうちに推移したと推定できる。ハプロルグープ O の下位群とはおもに弥生時代以降に流入したようであるが、以下、考古学的、歴史的側面からもこの考えを検証していく。
表2-9
→次頁「日本人のミトコンドリアDNAハプログループ」
〈文献〉
・Hammer et al. 2006. Dual origins of the Japanese: common ground for
hunter-gatherer and farmer Y chromosomes.
J Hum Genet. 2006;51(1):47-58.
Epub 2005 Nov 18.
・Shi,Hong , Hua Zhong, Yi Peng, Yong-Li Dong, Xue-Bin Qi, Feng Zhang, Lu-Fang Liu, Si-Jie Tan, Runlin Z Ma, Chun-Jie Xiao, R Spencer Wells, Li Jin, and Bing Su. 2008. Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations. BMC Biol. 2008; 6: 45. Published online 2008 October 29. doi: 10.1186/1741-7007-6-45
・Nonaka,I et al 2007, Y-chromosomal Binary Haplogroups in the Japanese Population and their Relationship to 16 Y-STR Polymorphisms
・Katoh,Toru 2004, Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis
・Yali Xue et al. 2013. Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times. Genetics.December 2013, 195 (4)
・Zegura SL1, Karafet TM, Zhivotovsky LA, Hammer MF.High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas.Mol Biol Evol. 2004 Jan;21(1):164-75. Epub 2003 Oct 31.