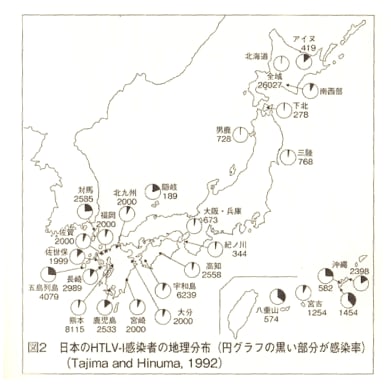
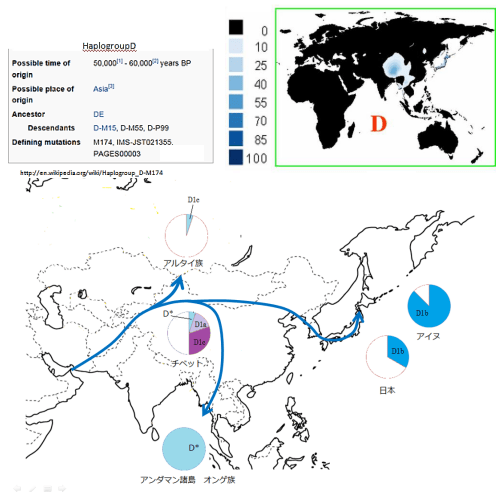
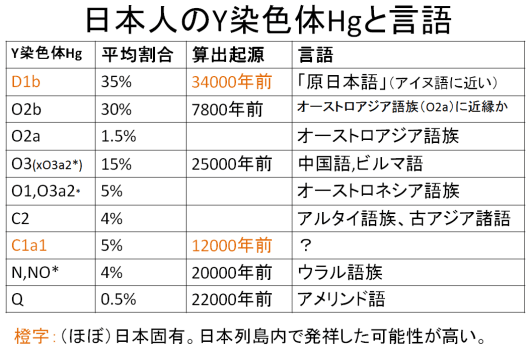
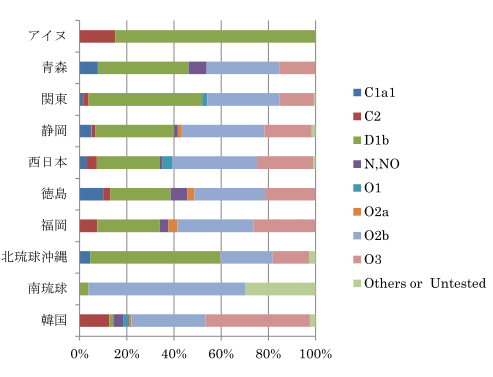
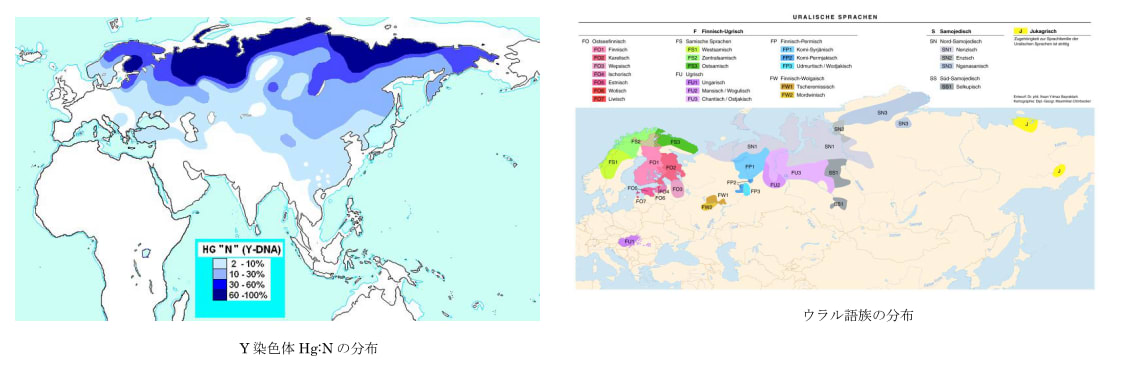
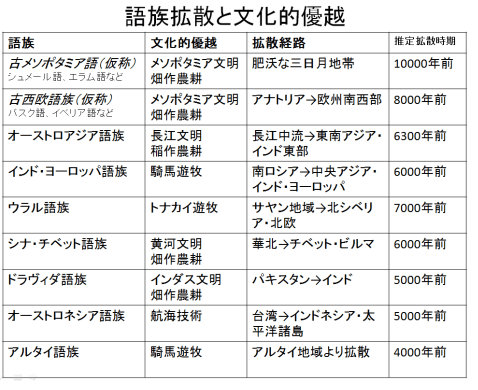
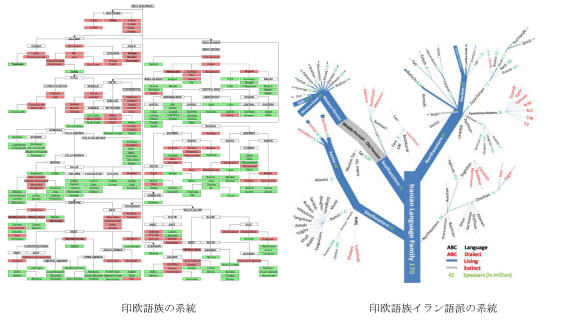
1990年代、日本人の起源を解き明かす画期的な研究成果が発表された。東京大学医学研究科の徳永勝士先生の研究グループが、HLAの多型により日本人の起源のかなりの部分を解き明かしたと発表したのである。
日本人の成り立ちについて述べる前に、まず最大のキーワードであるHLAについて、ある程度紹介しておこう。人類をはじめ高等動物が自分の体を守る仕組みである免疫系の働きは自己と非自己を区別することにある。即ち外から入ってきた病原微生物やウイルスなどの感染を受けて「変化した自己」の細胞を「正常な自己」の細胞と区別して攻撃し排除せねばならない。その識別のための「目印」として使われる分子グループを、人間の場合HLA(ヒト白血球抗原Human Leucocyte Antigen)と呼んでいる。
このような役割を果たすために、HLAの遺伝子群は複雑で多様性に富んでいる。HLAは第六染色体の短腕の上に密集して存在する一群の遺伝子から構成され、しかもそれぞれの遺伝子が著しい個人差を示す。よって人類集団のより詳細な研究に役立つのである。下図のようにHLAの遺伝子群は染色体上で近接しておりそれぞれのHLA遺伝子座の特定の対立遺伝子がセットを組んで親から子へと伝えられていく。このHLA遺伝子セットを「ハプロタイプ」とよびぶ。ハプロタイプは個人差だけでなく、著しい集団差があることでも知られる。これは他の遺伝子標識と比べ明らかな特徴である。

図2-1、図2-2(徳永1995)
このハプログループは非常に複雑なので全く同じものが異なる集団で別々に形成され、頻度を増すことは殆ど考えられない。よって同じハプロタイプが異なる集団で観察されれば、彼らは祖先集団を少なくとも一部は共有していると断定できる。またHLAハプロタイプは「保存性がいい」標識でもある。HLA遺伝子群は染色体上に密接に連鎖しており、いったん集団中で頻度を増したハプロタイプは数千年から数万年は存続することが期待されるからである。HLA遺伝子のセットの分布には明瞭な地域差・集団差が認められる。よって、容易に先祖集団の共通性(故郷)などを判断できるのである。
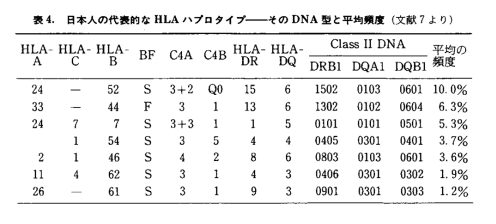
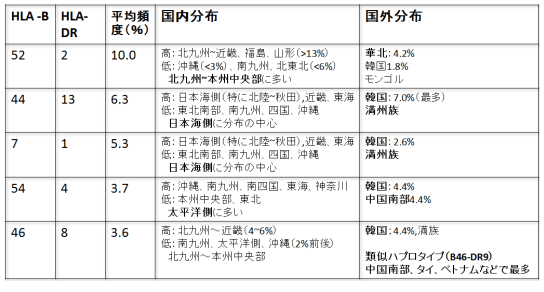
さて日本人におけるハプロタイプの上位8種類を示したのが下表である。徳永氏ほか多くの調査で各地に住む600人以上の日本人家族および東アジアの集団からハプロタイプデータを集めた。その結果、日本国内でも明らかな地域差が存在することがわかった。ここでは上位5型についてみてみよう。(以下徳永(1995)を引用)
表2-1 (徳永1995)

日本人全体で、最も多いハプロタイプB52-DR2(A24 -B 52-DR2-DQ1。以下B52-DR2)は北九州、山陽から近畿地方にかけて、そして山形や福島でも12~13%以上と高かった。対照的に南九州と東北地方の青森、岩手では6%前後、沖縄では2%程度と低かった。従ってこのハプログループは北九州から本州の西部や中央部にかけて多いタイプといえよう。日本の周辺部をみると、中国南部や東南アジアの諸民族には観察されず、対照的に中国北部の漢民族や韓国人で2%前後あることがわかった。さらに興味深いことに、これはモンゴル人においても最も多いハプロタイプである可能性が高いのに対し、中国東北地方の満族やオロチョン族などではまれで、シベリアのヤクートやバイカル湖のほとりのブリアートにおいてもほとんどみいだされていない。
2番目のハプログループ(B44-DR13)は、特に北陸地方から秋田にかけて多いことが知られており、また近畿や東海地方でも6%以上と多かった。対照的に東北地方の太平洋側や南九州、四国、沖縄では少ない傾向にあった。先に述べた一番多いハプロタイプ(B52-DR2)の分布状況と似ているが、日本海沿岸、とくに北陸地方に分布の中心がある点でいくらか異なっている。日本の周辺における分布をみても、先のハプロタイプ同様、中国南部よりは北部に多いハプログループであるが、詳しく見てみると違いが認められる。じつはこのハプログループは韓国人では7%以上と最も高頻度で見出されるタイプであり、また中国東北地方の満族にもかなりの頻度で見られた。にもかかわらず中国北部の漢民族やモンゴル人では殆ど見られなかった。表に示される3番目のハプログループ(B7-DR1)も同様な分布パターンを示していた。
4番目のハプログループ(B54-DR4)は、先の三つのハプロタイプとは対照的に、南方に分布が偏っているようにみえる。日本列島においては、特に沖縄で最も多いハプロタイプであり、南九州、南四国や東海地方、神奈川でも比較的多く観察された。一方、本州中央部や東北地方では頻度は低い傾向にあった。つまり、このハプロタイプは日本の南部から太平洋側にかけて多いハプロタイプといえよう。日本の周辺地域について見てみると、北方では韓国のみに多く観察された。興味深いことに、中国南部にすむいろいろな民族ではほとんど見られず、南方のタイ人やベトナム人でもまれであった。
さて、5番目のハプロタイプ(B46-DR8)であるが、その日本列島における分布は再び前二者に類似したパタンをとっていた。このハプログループは北九州から山陽、近畿地方にかけて4~6%と多い傾向にあり、沖縄、南九州や太平洋沿岸地域では2%前後と少ない。日本周辺部では韓国人と中国東北部の満族のみで多く、中国内でもその他の民族やシベリアあるいは東南アジアの集団には見られなかった。ところが、図12に示したように、このハプロタイプに非常に類似した別のハプロタイプ(B46-DR9)があって、中国南部、四川省やタイ、ベトナムの集団で最も多いことが知られている。この2つのハプロタイプは共通の祖先ハプロタイプに由来すると考えられるので、この両者が南方型と北方型という対照的な分布パタンをとっていることは非常に興味深い。
以上の例をみても、東アジアのHLAハプロタイプの分布状況から、はっきりとした地域差あるいは民族差が浮かび上がってくる。比較的均質と考えられてきた日本人の中にも明瞭な地域差が認められるわけである。(以上引用)
徳永氏の研究結果をまとめてみたのが下の表2-2である。
表2-2
(以下再び引用・多少改変)
HLA遺伝子及びハプロタイプの分布から、東アジアの諸集団、特に日本人の成立過程について考察したい。今まで述べてきたようにHLAハプロタイプの分布は明らかに異なっている。しかも、それぞれのハプロタイプがばらばらに分布しているのではなく線上のつながりを示し、少なくとも3つから4つの異なるパタンがみられる。この現象を説明する最も単純なシナリオは、日本人祖先が少なくとも3つから4つあり、それぞれ異なったルートから日本列島にやってきたというものであろう。
第1に、B52-DR2を高頻度にもつ集団が、中国北方より朝鮮半島を経て北九州や近畿地方に移住してきた可能性が考えられる。この祖先集団と現在のモンゴル高原周辺に住む人々との関係に多いに興味をそそられる。
第2に、B44-D13とB7-DR1で特徴づけられる集団が朝鮮半島あるいはその近隣を起点として、北陸地方などの日本海側に至った可能性が考えられる。先に述べたように、このハプロタイプをもっとも濃密にもつ集団は韓国人や中国の満族であったと考えられる。他のハプロタイプにも同様の分布パタンをとるものがあるので、以上2つの流れが現代日本人の遺伝的特徴に与えた大きな影響が想定される。
第3には、B54-DR4を多くもつ集団が、中国南部を起点として西南諸島や、九州、四国を経て、本州の太平洋岸に達するルートが考えられる。(ただしこのハプログループはおそらく海を渡って朝鮮半島にも伝わったのであろう。)
最後に、B46-DR8をもつ集団が考えられる。先に述べたように、おそらく祖先を共通する類似したハプロタイプ(B46-DR9)はあきらかに中国南部に由来するが、これらをもつグループは朝鮮半島を経由するかあるいは直接に、南九州ではなく北九州に到達したのかもしれない。
現時点でははっきりと断定できないが、以上のHLAハプロタイプとは別に、頻度は低いながら南九州と東北地方北部に共通して存在し、その間の本州中央部には極めて稀なHLAハプロタイプが観察されている。形質人類学の多くの成果も考え合わせると、これらが縄文時代人の特徴の一部を反映し、先に述べたハプログループはむしろ弥生時代に渡来してきた人々を特徴づけているのかもしれない。(以上引用)
なおアイヌ人のHLAハプロタイプは詳細研究が進んでいないが、日本人と異なったタイプが多いという。
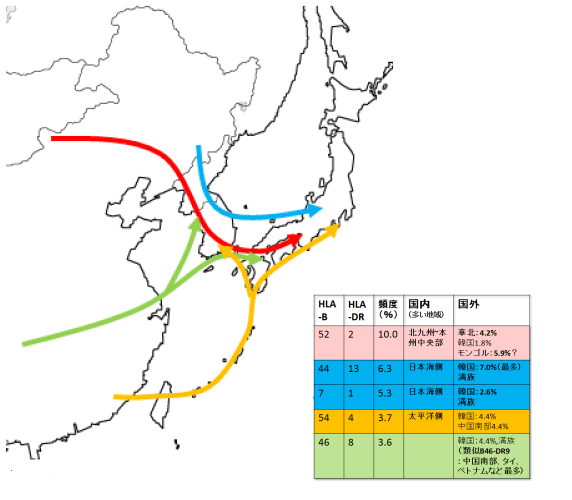
図2-5、表2-3
以上まとめてみると、大きく以下の4つの流れが認められる。
1. 中国大陸北部から朝鮮半島を経て北九州・近畿へ(赤)
2. 満州・朝鮮半島東部から日本海沿岸へ (青)
3. 中国南部から琉球諸島を経て太平洋側へ(オレンジ)
4. 中国大陸南部から直接、あるいは朝鮮半島を経由して北九州へ(緑)
さらにこれとは別に縄文系と想定される別の複数のハプロタイプが南九州や北東北に存在する。
(詳細は徳永(1995,1998,2003,2006)を参照)
このように、日本人は少なくとも5種類以上の集団から成り立っていることが分かったのである。ではそれぞれの集団がどの民族に近く、どのような言語を話し、いつ日本にやってきたのであろうか?
→次頁「日本人のY染色体Hg」
<文献>
徳永勝士 (1995)「HLA 遺伝子群からみた日本人のなりたち」『モンゴロイドの地球(3)日本人のなりたち』東京大学出版会,第 4 章,遺伝子からみた日本人,p193-210
徳永勝士 (1996) 「HLA の人類遺伝学」『日本臨床免疫学会会誌』=『Japanese journal of clinical immunology』19(6), 541-543
徳永勝士 (2003)「HLA と人類の移動」『Science of humanity Bensei 』(42), 4- 9, 東京:勉誠出版
徳永勝士 (2008)「HLA 遺伝子:弥生人には別ルートをたどってやってきた四つのグループがあった!」『日本人のルーツがわかる本』逆転の日本史編集部,東京:宝島社,p264-p280

















 図2-33、34
図2-33、34