ч
[編集] 遺伝子変換 (gene conversion)
V(D)J遺伝子再構成を終えた可変領域遺伝子が、V遺伝子上流に存在する偽遺伝子にランダムに置換されて、多様性をつくる。これを遺伝子変換 (gene conversion; GC) といい、主にニワトリでみられる[14][15]。1986年レイノーらにより報告された[16][17]
[編集] クラススイッチ組み換え (class switch recombination; CSR)
V(D)J遺伝子再構成等の過程を経て生まれたB細胞は、抗原の刺激を受けると成熟化し、増殖する。この際、重鎖定常領域 (CH) をコードする遺伝子にDNA改変が起こり、最初IgMを分泌していたB細胞はIgG等他のクラスの免疫グロブリンを産生する。同じ可変領域を異なる定常領域と組み合わせることにより、さらに多様な抗体を作り出す。このことをクラススイッチ組み換えという[11]。
[編集] 抗体医薬
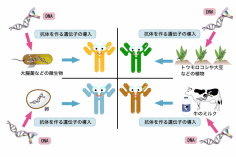
近年、モノクローナル抗体の持つ特異性を利用した医薬品の開発が進んでいる。抗体医薬は標的となる抗原に対して特異的に働くためにこれまでの医薬品よりも副作用を軽減させ、かつ高い治療効果が得られることが期待されている。2008年現在で関節リウマチ治療薬として抗TNF-α抗体であるインフリキシマブや抗IL-6抗体であるトシリズマブ、癌遺伝子HER2に対する抗体であるトラスツズマブなどがすでに臨床において使用されている。
[編集] 関連項目
ウィキメディア・コモンズには、抗体に関連するカテゴリがあります。
モノクローナル抗体
ポリクローナル抗体
抗原抗体反応
エピトープ
ワクチン
ELISA
ELISPOT
移行抗体
[編集] 参考文献
^ ラクダ科の動物は、重鎖だけで構成されるサイズの小さな抗体(ナノ抗体)を持つことが知られている。Hamers-Casterman, C. et al., "Naturally occurring antibodies devoid of light chains",Nature 363, 446-448 (1993).doi:10.1038/363446a0、W.W.ギブズ, 「開発進むナノ抗体医薬」,日経サイエンス2006年1月号
^ Porter RR. "The hydrolysis of rabbit γ-globulin and antibodies with crystalline papain."Biochemical Journal, 73, 1959, p.p. 119-127.PMID 14434282
^ Nisonoff A, Wissler FC, Lipman LN, Woernley DL. "Separation of univalent fragments from the bivalent rabbit antibody molecule by reduction of disulfide bonds."Archives of biochemistry and biophysics, 89, 1960, p.p. 230-44.PMID 14427334
^ Stavnezer J, Amemiya CT. "Evolution of isotype switching"Semin. Immunol.16, 2006, p.p. 257-275.PMID 15522624
^ Silverstein AM. "Paul Ehrlich's passion: the origins of his receptor immunology."Cellular Immunology, 194, 1999, p.p. 213-221.PMID 10383824
^ Edelman GM. "Journal of American Chemical Society", 81, 1959, p.p. 3155.
^ Edelman GM, Poulik MD. "Studies on structural units of the gamma-globulins."Journal of Experimental Medicine, 113, 1961, p.p. 861-884.PMID 13725659
^ Burnet FM. "A modification of Jerne's theory of antibody production using the concept of clonal selection."Australian Journal of Science, 20, 1957, p.p. 67-69.
^a b Hozumi N, Tonegawa S. "Evidence for somatic rearrangement of immunoglobulin genes coding for variable and constant regions."Proceedings of National Academy of Science of United States of America, 73, 1976, p.p. 3628-3632.PMID 824647
^ Tonegawa S. "Somatic generation of antibody diversity."Nature, 302, 1983, p.p. 575-581.PMID 6300689
^a b c Li Z, Woo CJ, Iglesias-Ussel MD, et al. "The generation of antibody diversity throuth somatic hypermutation and class switch recombination."Gene & Development, 18, 2004, p.p. 1-11.PMID 14724175
^ Market E, Papavasiliou FN."V(D)J recombination and the evolution of the adaptive immune system." PloS Biology, 1, 2003, p.p. 24-27.PMID 14551913
^ Rajewsky K, Forster I, Cumano A. "Evolutionary and somatic selection of the antibody repertoire in the mouse."Science, 238, 1987, p.p. 1088-1094.PMID 3317826
^ Weill JC, Reynaud CA, Lassila O, Pink JR. "Rearrangement of chicken immunoglobulin genes is not an ongoing process in the embryonic bursa of Fabricius."Proceedings of National Academy of Science of United States of America, 83, 1986, p.p. 3336-3340.PMID 3010290
^ Weill JC, Reynaud CA. "Rearrangement/hypermutation/gene conversion: When, where and why?"Immunology Today, 17, 1996, p.p. 92 -97.PMID 8808057
^ Reynaud CA, Anquez V, Dahan A, Weill JC. "A single rearrengement event generates most of the chicken immunoglobulin light chain diversity."Cell, 40, 1985, p.p. 283-291.PMID 3917859
^ "系統看護学講座 専門基礎① 解剖生理学 人体の構造と機能[1]"医学書院, p.p. 435.
この項目は、生物学に関連した書きかけの項目です。この記事を加筆・訂正などしてくださる協力者を求めています(プロジェクト:生命科学/Portal:生物学)。
この項目は、医学に関連した書きかけの項目です。この記事を加筆・訂正などしてくださる協力者を求めています(プロジェクト:医学/Portal:医学と医療)。
「http://ja.wikipedia.org/w/index.php?title=抗体&oldid=46385604」から取得
カテゴリ:
免疫学
タンパク質
抗体
隠しカテゴリ:
生物学関連のスタブ項目
医学関連のスタブ項目