








昆虫の呼吸―その5
気管-毛細気管呼吸
気管呼吸はシルル紀前期(4.4億年前)の頃にムカデやヤスデなどの多足類が初めて獲得して陸上に進出し、それが昆虫に受け継がれたと考えられています。
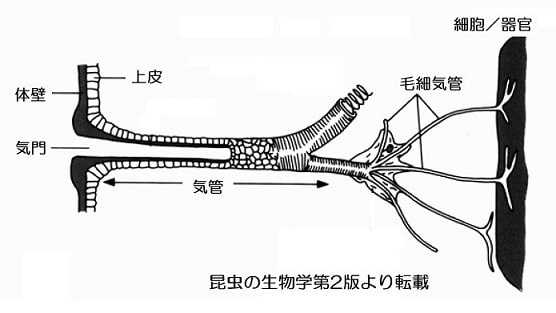
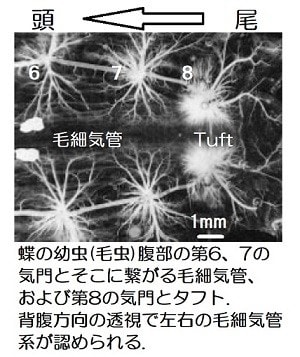
「昆虫の呼吸-3」で話題としたように、昆虫では気門を通って直径100ミクロン(μm)(0.1mm)~2μm程度の気管から多数に枝分かれした1~0.1μm(1万分の1mm)の毛細気管を通して酸素を体中の組織や細胞に送っています(駒井1996)。
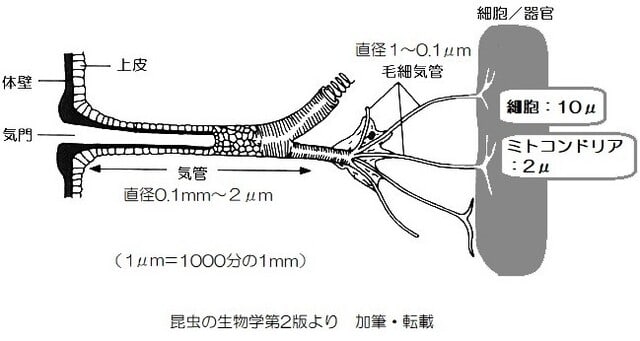
細胞で酸素が消費されると、気門付近の酸素濃度の高い空気から末梢の低酸素の毛細気管内へ酸素が供給されます。そのガス交換は、気管-毛細気管でおきる撹拌と拡散によるものです。
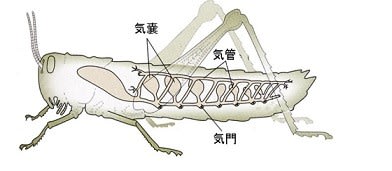
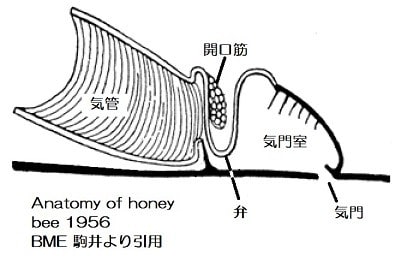
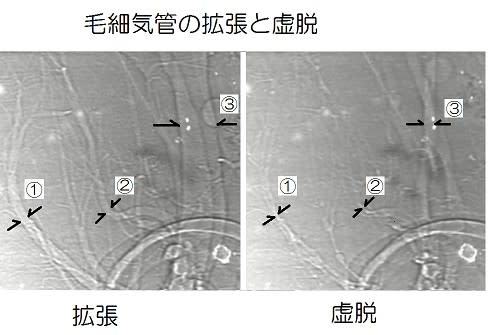
① 撹拌:飛翔筋など移動に使う筋肉が緊張すると気管系が圧迫されて管内の空気が排気される。その後に筋肉が弛緩すると拡張により新鮮な空気を吸い込むという気管系の換気運動(Weis-Fogh1964、Westneat 2003)によって古い空気の一部が新鮮な空気と入れ替わります。これについては「昆虫の呼吸-3」に写真を掲載し、動画の紹介をしたので参照してください。
こ例外にも昆虫の腹部は常に伸び縮みして気管内の空気の撹拌をしている。
② 拡散:撹拌に加えて、主に毛細気管系ではガス分子(酸素、二酸化炭素、水蒸気)がその濃度を均一にするように拡散していきます。末梢の毛細気管の酸素が細胞や組織で消費されて濃度が下がるとそれを補うように気管の酸素分子が拡散してきます。二酸化炭素と水蒸気は細胞から排出されると酸素とは逆に気管から気門に向かって拡散していきます。
ここでは、特に拡散について概説します
分子の拡散の性質
(以下では10の自乗を10^2、n乗を10^nなどと表しています)
空気中の酸素と窒素の分子は1ml中に合計で約2.7x10^19個(270億の10億倍)あって、その分子間距離は約3.4nm(ナノメートル:千分の1μm)と平均の分子直径0.38nmの10倍ほどです。このように小さくて密集している分子は平均速度400~500m/秒(時速1800km)で飛び回って、お互いに毎秒数百億回も衝突して散乱しています。
膨大な数の分子同士が衝突して散乱するために濃度や圧力が平均化されて均一になります。この現象が拡散です。
分子の数が多いために速度が大きくても次の衝突までに進める距離は短くて、酸素分子も窒素分子も約0.07μm(70nm)進むと次の分子と衝突します。この距離を平均自由行程といいます。
また、この距離を進む時間(平均自由時間)は100億分の1秒程度です。
目に見える太さの管や昆虫の気管では、この分子の拡散が十分に行われて濃度が均一になります。例えば管の一方の端Aで酸素の濃度が高く、反対側Bで窒素濃度が高かったとすると、酸素分子はAからBへ、窒素分子はBからAにそれぞれ拡散して管の中の酸素と窒素の濃度は均一になります。
しかし、毛細気管の直径:0.1~0.2μmのように、平均自由行程の0.07μmに近い極めて細い管の中では分子間の衝突よりも壁との間の衝突が増える結果、分子同士の衝突によって起きる拡散が障害されます。この毛細気管内で酸素が細胞に取り込まれて濃度が低下したときに拡散による酸素の供給速度が低下します。

毛細気管の最小直径が0.1~0.2μm程度なのは、この様な拡散の性質が原因となっているといわれています。
毛細気管の最小径0.1μmに対して酸素分子の大きさはその千分の1程度と極めて小さいので酸素分子は自由に流れるように思われますが、膨大な数の気体分子の激しい衝突と散乱が拡散現象の本質であるために、管壁への衝突が大きくなると分子の移動が障害されることになります。
参考文献
駒井豊 総説. 昆虫のガス交換機序BME11:19-28. 1996
本川達夫 ゾウの時間ネズミの時間 中公新書 1992
松香光夫ほか 昆虫の生物学 第2版 玉川大学出版 1992
Weis-Fogh T. J Exp Biol 41: 229-56, 1964
Westneat,MW. Science vol299 558-560 2003
Hetz, S.K. Nature. 433: 516-519. 2005.
気管-毛細気管呼吸
気管呼吸はシルル紀前期(4.4億年前)の頃にムカデやヤスデなどの多足類が初めて獲得して陸上に進出し、それが昆虫に受け継がれたと考えられています。
「昆虫の呼吸-3」で話題としたように、昆虫では気門を通って直径100ミクロン(μm)(0.1mm)~2μm程度の気管から多数に枝分かれした1~0.1μm(1万分の1mm)の毛細気管を通して酸素を体中の組織や細胞に送っています(駒井1996)。
細胞で酸素が消費されると、気門付近の酸素濃度の高い空気から末梢の低酸素の毛細気管内へ酸素が供給されます。そのガス交換は、気管-毛細気管でおきる撹拌と拡散によるものです。
① 撹拌:飛翔筋など移動に使う筋肉が緊張すると気管系が圧迫されて管内の空気が排気される。その後に筋肉が弛緩すると拡張により新鮮な空気を吸い込むという気管系の換気運動(Weis-Fogh1964、Westneat 2003)によって古い空気の一部が新鮮な空気と入れ替わります。これについては「昆虫の呼吸-3」に写真を掲載し、動画の紹介をしたので参照してください。
こ例外にも昆虫の腹部は常に伸び縮みして気管内の空気の撹拌をしている。
② 拡散:撹拌に加えて、主に毛細気管系ではガス分子(酸素、二酸化炭素、水蒸気)がその濃度を均一にするように拡散していきます。末梢の毛細気管の酸素が細胞や組織で消費されて濃度が下がるとそれを補うように気管の酸素分子が拡散してきます。二酸化炭素と水蒸気は細胞から排出されると酸素とは逆に気管から気門に向かって拡散していきます。
ここでは、特に拡散について概説します
分子の拡散の性質
(以下では10の自乗を10^2、n乗を10^nなどと表しています)
空気中の酸素と窒素の分子は1ml中に合計で約2.7x10^19個(270億の10億倍)あって、その分子間距離は約3.4nm(ナノメートル:千分の1μm)と平均の分子直径0.38nmの10倍ほどです。このように小さくて密集している分子は平均速度400~500m/秒(時速1800km)で飛び回って、お互いに毎秒数百億回も衝突して散乱しています。
膨大な数の分子同士が衝突して散乱するために濃度や圧力が平均化されて均一になります。この現象が拡散です。
分子の数が多いために速度が大きくても次の衝突までに進める距離は短くて、酸素分子も窒素分子も約0.07μm(70nm)進むと次の分子と衝突します。この距離を平均自由行程といいます。
また、この距離を進む時間(平均自由時間)は100億分の1秒程度です。
目に見える太さの管や昆虫の気管では、この分子の拡散が十分に行われて濃度が均一になります。例えば管の一方の端Aで酸素の濃度が高く、反対側Bで窒素濃度が高かったとすると、酸素分子はAからBへ、窒素分子はBからAにそれぞれ拡散して管の中の酸素と窒素の濃度は均一になります。
しかし、毛細気管の直径:0.1~0.2μmのように、平均自由行程の0.07μmに近い極めて細い管の中では分子間の衝突よりも壁との間の衝突が増える結果、分子同士の衝突によって起きる拡散が障害されます。この毛細気管内で酸素が細胞に取り込まれて濃度が低下したときに拡散による酸素の供給速度が低下します。
毛細気管の最小直径が0.1~0.2μm程度なのは、この様な拡散の性質が原因となっているといわれています。
毛細気管の最小径0.1μmに対して酸素分子の大きさはその千分の1程度と極めて小さいので酸素分子は自由に流れるように思われますが、膨大な数の気体分子の激しい衝突と散乱が拡散現象の本質であるために、管壁への衝突が大きくなると分子の移動が障害されることになります。
参考文献
駒井豊 総説. 昆虫のガス交換機序BME11:19-28. 1996
本川達夫 ゾウの時間ネズミの時間 中公新書 1992
松香光夫ほか 昆虫の生物学 第2版 玉川大学出版 1992
Weis-Fogh T. J Exp Biol 41: 229-56, 1964
Westneat,MW. Science vol299 558-560 2003
Hetz, S.K. Nature. 433: 516-519. 2005.
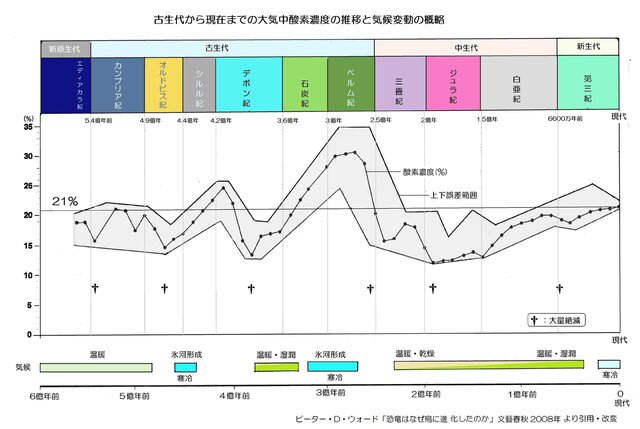

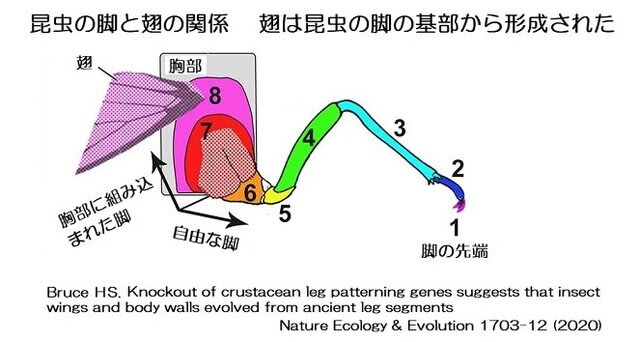



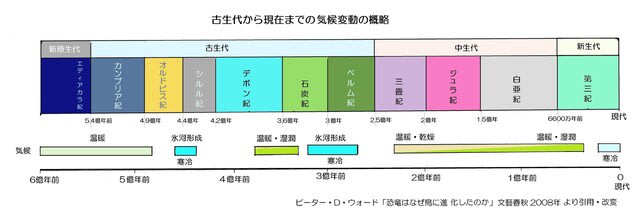 ・進化史の概略
・進化史の概略







