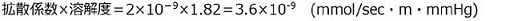両生類の呼吸―その7
代謝率について
******今回の要約****************************************************************
「両生類の呼吸-その4」では両生類の酸素消費量(代謝率)を有尾目サイレンで得られた結果を用いました。今回は有尾目、無尾目の広範な体重分布に対する代謝率のデータから15℃、20℃、25℃の体重―酸素消費量の関係式(回帰式)を求めました。
その回帰式を使って「両生類の呼吸-その4」と同様にして測定温度20℃で体重13gのカエルの酸素消費量と皮膚からの酸素消費量を比較しました。表皮の厚さは文献から0.05mmとしました。
その結果、20℃の陸上にいる13gのカエルの安静時酸素消費量は0.0135ml/分であり皮膚からの酸素吸収量は0.0183ml/分となりました。
皮膚酸素吸収量が酸素消費量よりも十分に大きいので、肺呼吸を併用すれば陸上での運動時でも低酸素になることはないと推測されました。
*********************************************************************************
「両生類の呼吸-その4」では、両生類の代謝率については、シュミットニールセンの教科書に記載された水棲の有尾目サイレン科で測定された関係式を両生類全般に使うことにして、代謝率P(kcal/日)と体重M(㎏)の関係式をP=0.79M^0.65 としました。(M^0.65はMの0.65乗を示します。以下同様)
けれども、その後いくつかの文献と書籍から、両生類のうち有尾目(サンショウウオ等)、無尾目(カエル)の体重と酸素消費量(代謝率)の関係式を求めることができました。
文献などのデータは体重をkg、酸素消費量を ml/分に変換してExcel上で両対数軸の散布図から累乗近似曲線により関係式(回帰式)求めました。その散布図は最後にまとめて記載しています。
また、後述するように、動物では1mlの酸素を代謝に利用すると4.8calの発熱量があることを使って酸素消費量を熱量(kcal)に換算して、は虫類、哺乳類と比較します。
1.有尾目の代謝率:サイレンとその他有尾目
〇サイレン(sirenidae)
上記のサイレンの関係式はUltsch(1974)の測定によるものです。彼は平均体重0.36gから1310gの水棲の有尾目サイレンを酸素で飽和された25℃の水中に2時間潜水させてから、1時間以上に亘り水中の酸素分圧の低下を測定して酸素消費量の平均値を求めました。
回帰式は、W(酸素消費量)=0.1113M^0.654(ml/分, M:kg)、相関係数は0.9896でした。(GR Ultsch. Gas exchange and metabolism in the sirenidae1974表9より)
○有尾目(サイレンを除く)と無尾目の回帰式
教科書(Environmental Physiology of the Amphibians Table12.1)には1897年から1989年までの間に報告された約750件の両生類の酸素消費量の測定値が一覧されています。
その表では、有尾目、無尾目、無足目に対して、5℃~30℃の範囲で体重(g)、酸素消費量(ml/h)、固有酸素消費量(ml/g/h)の測定値を利用できます。
サイレンを除く有尾目(サンショウウオ)と無尾目(カエル)について15℃、20℃、25℃の体重(g)、酸素消費量(ml/h)をKgとml/分に変換して回帰式を求め、以下の表にまとめました。
サイレンについてはUltsch(1974)のデータです。無足目についてはデータ量が少ないので回帰式を求めていません
なお、サイレンを除く有尾目と無尾目の回帰式を求めるときに、回帰式の係数と指数が大きく変化しない範囲で、散布図から4~5つの著しい外れ値を除いて相関係数を大きくしています。

・これらの回帰式の相関係数がどれも0.8以上で体重と酸素消費量の間に強い相関がみられます。
・回帰式の指数はサイレンでは0.65だが、有尾類、無尾類ではどの温度でも0.8程度です。有尾類、無尾類では多くの種が混在したデータから回帰式が求められていることが、サイレン1種の回帰式との違いの原因かもしれません。指数については、イモリ単独あるいはアマガエル単独の広範囲のデータから得られた回帰式の検討が必要かもしれません。
・有尾目と無尾目の温度変化では15℃と20℃の係数に大きな変化はないが、25℃になる と約2倍になって、体重を同じとすれば酸素消費量は2倍です。
両生類はもともと冷涼な15~20℃の環境に適応していて25℃では“暑すぎる”ために酸素消費量が増えているのでしょうか。
両生類やは虫類のような外温動物では温度変化により細胞内の代謝速度が変わります。一般的に代謝量の変化をみる指標としてある温度T℃とそれより10℃高いT+10℃の酸素消費量の比Q10 が用いられていて、Q 10の値は一般に2~3の間です。
有尾類と無尾類で15℃から25℃で酸素消費量が2倍になっているのはQ10=2に相当していて、この回帰式が代謝を反映していると考えられます。
○カエルの酸素消費量(代謝率)と皮膚からの酸素吸収量の再検討
「両生類の呼吸―その4」ではサイレンの回帰式を使って、25℃で体重1kgのカエルの酸素吸収量と代謝量をみましたが、ここでは今回得られた回帰式と平均体重を用い、表皮の厚さを再検討して酸素消費量と皮膚の酸素吸収量をみてみます。
まず、消費量はカエル20℃の式: W=0.476xM^0.82、体重Mは中央値の13g。
酸素消費量は、 W=0.476x(0.013)^0.82=0.0135ml/分となります。
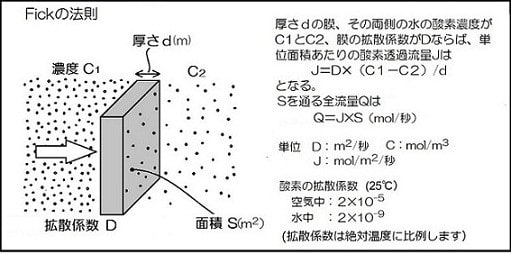
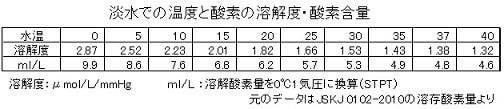
また、カエルの表皮の厚さについてはヒトに近い0.3mmと仮定しましたが、今回文献を渉猟したところ、1951年の沢近氏の蛙皮についての論文が見つかりました。それによると、トノサマカエルの表皮の厚みは季節で変化し、背と腹でも差がみられて、平均37µ(概ね20~60µの範囲)とのことです。今回は表皮の厚さとして誤差も含めて50µ(0.05mm)と前回の1/6としてして計算します。
間質の酸素拡散定数は1.1x10^(-9)mmol/sec/m/mmHg(20℃)と前回と同様です。
表皮の厚さ0.05mmとして前回と同様に計算すると、皮膚からの酸素吸収量は単位表面積当たり3.17x10^(-4)ml /分です。
カエルの体表面積S(cm^2)と体重M(kg)の回帰式は前回と同じで:S=1060xM^(0.67)
これより、13gのカエルの体表面積=1060x0.013^0.67=57.8 cm^2なので、
吸収酸素量=3.17x10^(-4)x57.8=0.0183ml/分 です。
以上から、20℃の陸上にいる13gのカエルは安静時に酸素の消費量:0.0135ml/分であり皮膚からの酸素吸収量は0.0183ml/分となりました。
皮膚酸素吸収量が酸素消費量よりも十分に大きいので、肺呼吸を併用すれば陸上での運動時に低酸素となることはないでしょう。また空気に良くさらされた冷涼な水中ならば十分に酸素を取り入れることができると推測されました。
2.爬虫類・哺乳類の代謝式
爬虫類の代謝率も多様です。いくつかの文献の回帰式を表にしました。
回帰式の係数は1前後であり、体重Mの指数は0.8前後でした。係数の値から爬虫類の酸素消費量は15~20℃の環境中の両生類の5倍程度でした。
両生類ではこの係数に相当するのは25℃のカエルの回帰式の係数でした。爬虫類の測定温度は30℃と両生類よりも高いことが関係しているのでしょう。

3.哺乳類の代謝式
哺乳類では体重M(kg)と酸素消費量W(ml/分)の回帰式は
○ W=11.3M^0.75 (シュミットニールセン)
教科書中の単位では体重M(kg)と熱量P(kcal/日):P=77.9M^0.75です。
(酸素1mlで4.8calとして換算)
○ W=12.2M^0.75 (本川 ゾウの時間ネズミの時間)
本文中の単位では体重M(kg)とW(ワット):W=4.1M^0.75です。
(1Watt=1/4.19 cal/secで換算)
哺乳類は両生類や爬虫類と違って外気温に依存しない内温動物なので体温を36~38℃に保つために、エネルギー消費量(代謝率)は大きくなっています。
回帰式の係数で比較すると、爬虫類の代謝率は15℃の有尾目の約5倍、哺乳類は30℃の爬虫類の約10倍の代謝率になっていました。
今回は、多くの文献や教科書から引用したデータで両生類の代謝率を算出し、それと爬虫類、哺乳類の代謝率を比較するためにずいぶん時間がかかりました。データは全てが最新の信頼性の高い値ばかりではないことや、データ処理における誤差も含まれていると思いますが、それぞれの生息環境下での代謝率は、おおむね哺乳類の1/10が爬虫類、爬虫類の1/5が両生類との結果でした。
今回の結果は「両生類の呼吸-その4」よりも正確と思われます。しかし両生類の動物種の間で間質の酸素拡散定数を同一としていますが、動物種毎の拡散定数は測定されていないのでしょうか?
<酸素消費量と熱量(カロリー)の関係について>
酸素消費量から熱量のカロリーへの変換については次のように考えます。
動物が代謝する炭水化物、脂質、タンパク質を代表する物質として、グルコース、パルミチン酸、アラニンを取り上げます。それぞれが酸素1Lで代謝されるときの熱量は、グルコースは21.4kJ(5.11kcal)、パルミチン酸は19.1kJ(4.56kcal)、アラニンは19.6kJ(4.65kcal) とほぼ20kJ程度です。動物によりこれらの代謝割合が変わっても10%以内(4.5~5.1kcal)なので、平均的に動物では酸素1Lの消費によって20.1kJ (4.8kcal)の熱量が産生するとしています(Dejours. P. 呼吸生理学の基礎 表2-1、およびK. Schmidt-Nielsen動物生理学:エネルギー代謝から引用)。
つまり動物においては、消費酸素量1ml当たり4.8calと換算されます。これを用いれば、酸素消費量を動物の代謝熱量に換算できます。
参考文献
1. K. Schmidt-Nielsen スケーリング: 動物設計論 コロナ社 東京1998
2..K. Schmidt-Nielsen 動物生理学 東京大学出版会 東京 2007
3 .Dejours P. 呼吸生理学の基礎 真興交易医書出版部 東京 1983
4 .GR Ultsch. Gas exchange and metabolism in the sirenidae (Amphibia: Caudata). Comparative Biochemistry and Physiology Part A: Physiology 47;p485-498 1974,
5. Feder ME. Environmental Physiology of the Amphibians. p317-329 table 12.1 Univ. of Chicago press. Chicago1992.
6. 沢近 巽 蛙皮の季節的変化、特にその組織測定. Archivum histologicum japonicum 2:79-83 1951
7 .LK Maxwell et,al. Intraspecific allometry of standard metabolic rate in green iguanas, Iguana iguana CBP Part A 136: 301–310 2003
8 .Seymour RS et,al. Scaling of standard metabolic rate in estuarine crocodiles Crocodylus porosus JCP-B 183:491-500 2013
9 .RM. Andrews, FH Pough Metabolism of Squamate Reptiles: Allometric and Ecological Relationships Physiological Zoology, 58: 214-231 1985



代謝率について
******今回の要約****************************************************************
「両生類の呼吸-その4」では両生類の酸素消費量(代謝率)を有尾目サイレンで得られた結果を用いました。今回は有尾目、無尾目の広範な体重分布に対する代謝率のデータから15℃、20℃、25℃の体重―酸素消費量の関係式(回帰式)を求めました。
その回帰式を使って「両生類の呼吸-その4」と同様にして測定温度20℃で体重13gのカエルの酸素消費量と皮膚からの酸素消費量を比較しました。表皮の厚さは文献から0.05mmとしました。
その結果、20℃の陸上にいる13gのカエルの安静時酸素消費量は0.0135ml/分であり皮膚からの酸素吸収量は0.0183ml/分となりました。
皮膚酸素吸収量が酸素消費量よりも十分に大きいので、肺呼吸を併用すれば陸上での運動時でも低酸素になることはないと推測されました。
*********************************************************************************
「両生類の呼吸-その4」では、両生類の代謝率については、シュミットニールセンの教科書に記載された水棲の有尾目サイレン科で測定された関係式を両生類全般に使うことにして、代謝率P(kcal/日)と体重M(㎏)の関係式をP=0.79M^0.65 としました。(M^0.65はMの0.65乗を示します。以下同様)
けれども、その後いくつかの文献と書籍から、両生類のうち有尾目(サンショウウオ等)、無尾目(カエル)の体重と酸素消費量(代謝率)の関係式を求めることができました。
文献などのデータは体重をkg、酸素消費量を ml/分に変換してExcel上で両対数軸の散布図から累乗近似曲線により関係式(回帰式)求めました。その散布図は最後にまとめて記載しています。
また、後述するように、動物では1mlの酸素を代謝に利用すると4.8calの発熱量があることを使って酸素消費量を熱量(kcal)に換算して、は虫類、哺乳類と比較します。
1.有尾目の代謝率:サイレンとその他有尾目
〇サイレン(sirenidae)
上記のサイレンの関係式はUltsch(1974)の測定によるものです。彼は平均体重0.36gから1310gの水棲の有尾目サイレンを酸素で飽和された25℃の水中に2時間潜水させてから、1時間以上に亘り水中の酸素分圧の低下を測定して酸素消費量の平均値を求めました。
回帰式は、W(酸素消費量)=0.1113M^0.654(ml/分, M:kg)、相関係数は0.9896でした。(GR Ultsch. Gas exchange and metabolism in the sirenidae1974表9より)
○有尾目(サイレンを除く)と無尾目の回帰式
教科書(Environmental Physiology of the Amphibians Table12.1)には1897年から1989年までの間に報告された約750件の両生類の酸素消費量の測定値が一覧されています。
その表では、有尾目、無尾目、無足目に対して、5℃~30℃の範囲で体重(g)、酸素消費量(ml/h)、固有酸素消費量(ml/g/h)の測定値を利用できます。
サイレンを除く有尾目(サンショウウオ)と無尾目(カエル)について15℃、20℃、25℃の体重(g)、酸素消費量(ml/h)をKgとml/分に変換して回帰式を求め、以下の表にまとめました。
サイレンについてはUltsch(1974)のデータです。無足目についてはデータ量が少ないので回帰式を求めていません
なお、サイレンを除く有尾目と無尾目の回帰式を求めるときに、回帰式の係数と指数が大きく変化しない範囲で、散布図から4~5つの著しい外れ値を除いて相関係数を大きくしています。
・これらの回帰式の相関係数がどれも0.8以上で体重と酸素消費量の間に強い相関がみられます。
・回帰式の指数はサイレンでは0.65だが、有尾類、無尾類ではどの温度でも0.8程度です。有尾類、無尾類では多くの種が混在したデータから回帰式が求められていることが、サイレン1種の回帰式との違いの原因かもしれません。指数については、イモリ単独あるいはアマガエル単独の広範囲のデータから得られた回帰式の検討が必要かもしれません。
・有尾目と無尾目の温度変化では15℃と20℃の係数に大きな変化はないが、25℃になる と約2倍になって、体重を同じとすれば酸素消費量は2倍です。
両生類はもともと冷涼な15~20℃の環境に適応していて25℃では“暑すぎる”ために酸素消費量が増えているのでしょうか。
両生類やは虫類のような外温動物では温度変化により細胞内の代謝速度が変わります。一般的に代謝量の変化をみる指標としてある温度T℃とそれより10℃高いT+10℃の酸素消費量の比Q10 が用いられていて、Q 10の値は一般に2~3の間です。
有尾類と無尾類で15℃から25℃で酸素消費量が2倍になっているのはQ10=2に相当していて、この回帰式が代謝を反映していると考えられます。
○カエルの酸素消費量(代謝率)と皮膚からの酸素吸収量の再検討
「両生類の呼吸―その4」ではサイレンの回帰式を使って、25℃で体重1kgのカエルの酸素吸収量と代謝量をみましたが、ここでは今回得られた回帰式と平均体重を用い、表皮の厚さを再検討して酸素消費量と皮膚の酸素吸収量をみてみます。
まず、消費量はカエル20℃の式: W=0.476xM^0.82、体重Mは中央値の13g。
酸素消費量は、 W=0.476x(0.013)^0.82=0.0135ml/分となります。
また、カエルの表皮の厚さについてはヒトに近い0.3mmと仮定しましたが、今回文献を渉猟したところ、1951年の沢近氏の蛙皮についての論文が見つかりました。それによると、トノサマカエルの表皮の厚みは季節で変化し、背と腹でも差がみられて、平均37µ(概ね20~60µの範囲)とのことです。今回は表皮の厚さとして誤差も含めて50µ(0.05mm)と前回の1/6としてして計算します。
間質の酸素拡散定数は1.1x10^(-9)mmol/sec/m/mmHg(20℃)と前回と同様です。
表皮の厚さ0.05mmとして前回と同様に計算すると、皮膚からの酸素吸収量は単位表面積当たり3.17x10^(-4)ml /分です。
カエルの体表面積S(cm^2)と体重M(kg)の回帰式は前回と同じで:S=1060xM^(0.67)
これより、13gのカエルの体表面積=1060x0.013^0.67=57.8 cm^2なので、
吸収酸素量=3.17x10^(-4)x57.8=0.0183ml/分 です。
以上から、20℃の陸上にいる13gのカエルは安静時に酸素の消費量:0.0135ml/分であり皮膚からの酸素吸収量は0.0183ml/分となりました。
皮膚酸素吸収量が酸素消費量よりも十分に大きいので、肺呼吸を併用すれば陸上での運動時に低酸素となることはないでしょう。また空気に良くさらされた冷涼な水中ならば十分に酸素を取り入れることができると推測されました。
2.爬虫類・哺乳類の代謝式
爬虫類の代謝率も多様です。いくつかの文献の回帰式を表にしました。
回帰式の係数は1前後であり、体重Mの指数は0.8前後でした。係数の値から爬虫類の酸素消費量は15~20℃の環境中の両生類の5倍程度でした。
両生類ではこの係数に相当するのは25℃のカエルの回帰式の係数でした。爬虫類の測定温度は30℃と両生類よりも高いことが関係しているのでしょう。
3.哺乳類の代謝式
哺乳類では体重M(kg)と酸素消費量W(ml/分)の回帰式は
○ W=11.3M^0.75 (シュミットニールセン)
教科書中の単位では体重M(kg)と熱量P(kcal/日):P=77.9M^0.75です。
(酸素1mlで4.8calとして換算)
○ W=12.2M^0.75 (本川 ゾウの時間ネズミの時間)
本文中の単位では体重M(kg)とW(ワット):W=4.1M^0.75です。
(1Watt=1/4.19 cal/secで換算)
哺乳類は両生類や爬虫類と違って外気温に依存しない内温動物なので体温を36~38℃に保つために、エネルギー消費量(代謝率)は大きくなっています。
回帰式の係数で比較すると、爬虫類の代謝率は15℃の有尾目の約5倍、哺乳類は30℃の爬虫類の約10倍の代謝率になっていました。
今回は、多くの文献や教科書から引用したデータで両生類の代謝率を算出し、それと爬虫類、哺乳類の代謝率を比較するためにずいぶん時間がかかりました。データは全てが最新の信頼性の高い値ばかりではないことや、データ処理における誤差も含まれていると思いますが、それぞれの生息環境下での代謝率は、おおむね哺乳類の1/10が爬虫類、爬虫類の1/5が両生類との結果でした。
今回の結果は「両生類の呼吸-その4」よりも正確と思われます。しかし両生類の動物種の間で間質の酸素拡散定数を同一としていますが、動物種毎の拡散定数は測定されていないのでしょうか?
<酸素消費量と熱量(カロリー)の関係について>
酸素消費量から熱量のカロリーへの変換については次のように考えます。
動物が代謝する炭水化物、脂質、タンパク質を代表する物質として、グルコース、パルミチン酸、アラニンを取り上げます。それぞれが酸素1Lで代謝されるときの熱量は、グルコースは21.4kJ(5.11kcal)、パルミチン酸は19.1kJ(4.56kcal)、アラニンは19.6kJ(4.65kcal) とほぼ20kJ程度です。動物によりこれらの代謝割合が変わっても10%以内(4.5~5.1kcal)なので、平均的に動物では酸素1Lの消費によって20.1kJ (4.8kcal)の熱量が産生するとしています(Dejours. P. 呼吸生理学の基礎 表2-1、およびK. Schmidt-Nielsen動物生理学:エネルギー代謝から引用)。
つまり動物においては、消費酸素量1ml当たり4.8calと換算されます。これを用いれば、酸素消費量を動物の代謝熱量に換算できます。
参考文献
1. K. Schmidt-Nielsen スケーリング: 動物設計論 コロナ社 東京1998
2..K. Schmidt-Nielsen 動物生理学 東京大学出版会 東京 2007
3 .Dejours P. 呼吸生理学の基礎 真興交易医書出版部 東京 1983
4 .GR Ultsch. Gas exchange and metabolism in the sirenidae (Amphibia: Caudata). Comparative Biochemistry and Physiology Part A: Physiology 47;p485-498 1974,
5. Feder ME. Environmental Physiology of the Amphibians. p317-329 table 12.1 Univ. of Chicago press. Chicago1992.
6. 沢近 巽 蛙皮の季節的変化、特にその組織測定. Archivum histologicum japonicum 2:79-83 1951
7 .LK Maxwell et,al. Intraspecific allometry of standard metabolic rate in green iguanas, Iguana iguana CBP Part A 136: 301–310 2003
8 .Seymour RS et,al. Scaling of standard metabolic rate in estuarine crocodiles Crocodylus porosus JCP-B 183:491-500 2013
9 .RM. Andrews, FH Pough Metabolism of Squamate Reptiles: Allometric and Ecological Relationships Physiological Zoology, 58: 214-231 1985