夏!! …といえば水遊びたろう。水泳・マリンスポーツ・釣りなんでもござれ。
……というわけで(???)、読者におかれましては、水辺の捕食者と聞かれて何を思い浮かべるだろうか?日本人なら、真っ先にヒグマか水鳥を挙げるだろう。あるいはペット人気の強いカワウソ ――本来ペットにすべきではないが―― や、荒くれ者のワニを連想する人もいるかもしれない。
それでは次に、今から1億年前の白亜紀へ思考を飛ばそう。そこで水辺の捕食者として振る舞っているのは、多種多様なワニ類とスピノサウルス類だ。どちらも細長い顎を持ち、かたや水中から、かたや岸辺から。次々と魚を仕留めて丸呑みにしていた。
さらに遡ろう。今から2億年以上に昔の三畳紀では、ワニそっくりの爬虫類が何種類かいたし、オオサンショウウオでさえ可愛く見える“化け物イモリ”もいた。もちろん主食は魚である。
……それでは、もっともっと遡って、今から3億年前。ようやく陸上で安定した生態系か形作られた時代へ向かってみよう。そこの水辺には誰が君臨していたのか……? まず目に付くのはヌルヌル這い回る両生類だ。これは三畳紀の化け物イモリの祖先筋にあたる。今でこそ意外だが、かつて両生類は水辺で最強の捕食者だったのだ(#1)。
がしかし、そこへ水飛沫とともに乱入した生物がいた。不届き千万、おのれ面見せぃ。
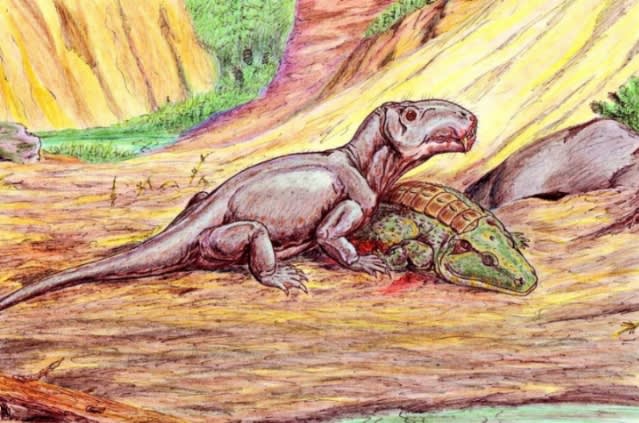
…顎は細長く、ズラリと並んだ歯は鋭い。四肢は短く頑丈で、胴体は細長い。一方チロリーんと伸びた尻尾は貧弱極まりない。…おそらく体表は鱗ないしシワだらけの硬い外皮に覆われていた。これこそが今回の主役、オフィアコドン《Ophiacodon》だ!!

(オフィアコドンの生態復元 wikiメディアより)
《基本情報》
学名→オフィアコドン《Ophiacodon》
全長→最大3.5メートル
食性→魚食(動きの遅い魚類や両生類)
生息時代・地域→約3億年前の北アメリカとヨーロッパの水辺(川や沼沢地)
先に述べておこう。
オフィアコドン(科)は爬虫類ではない。ワニやトカゲはおろか、あらゆる爬虫類とは無関係で、むしろ我々哺乳類に近い動物だ。かつては諸々をごった煮にして“哺乳類型爬虫類《mammal like reptiles》”と呼ばれていた時代もあったが、今では最基盤の単弓類と呼ばれている。
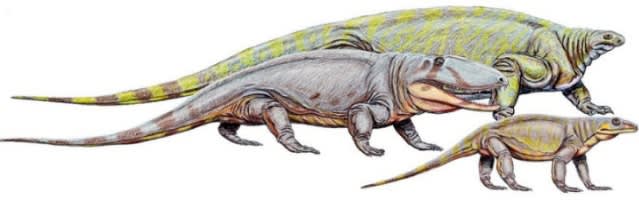
(最基盤の単弓類たち。奥がコティロリンクス、中がオフィアコドン、手前がヴァラノプス Wikiメディアより)
( ・ω・) 「あれ?“盤竜類《Pelycosauria》”っていう分類は違うのかい?」
今こんな質問が届いた。…良い質問だ。今から5年かそこら前まで、このグループ名は図鑑・専門書を問わず使われていた。有名なディメトロドンやエダフォサウルスなども所属していて、グループ名も彼らの背ビレから取られている。もちろん我らがオフィアコドンもその一員だった……。
がしかし、諸君朗報である(・∀・)
現在では盤竜類なる分類は空中分解してしまった。
エエッー!?) …このあたりは本筋とズレるため深堀りしないが、ともかく今後は“”を付けて表記するか、さもなくは素直に最基盤の単弓類と呼ぶべきだろう ――そのあたり便利な分類名が欲しいと思う今日このごろだ。

(いわゆる“盤竜類”の系統図。左に行くほど基盤的で、右に行くほど哺乳類に近い(派生的)である。頭骨の形態は実に様々だ (#C))
細かな分類はさておき、こうした最基盤の単弓類は、石炭紀の後半から本格的な多様化を始めた。というか、単弓類はフライング気味に進化のレースへ乗り出した。当時の爬虫類がどれも全長1mを超えない中、2m級の種類をポンポン量産していたのだ。…もっとも、それがあるグループの逆鱗に触れ、さらには地獄の釜の蓋を開いてしまったのだが ――それについては次回語りたい。
てなわけで、前振りはそこまで。ここからはオフィアコドン(科)の身体的特徴をまとめていきたい。ただし下半身からだ。
《尻尾》

(オフィアコドンの組み立て骨格。尻尾の貧弱さがよく分かる (#2))
情★報★不★足(´Д⊂ヽ
…実はオフィアコドン(に限らず初期の単弓類全般)の尻尾は資料が少ない。古来はチャールズ・ナイトの時代より、大根のごとき太ましい尻尾が描かれてきたのだが、ぶっちゃけ確たる証拠はない。
むしろ怪しいネットの吹き溜まりで手に入れた情報からするに、彼ら初期の単弓類の尻尾(尾椎)は、ちろーんと伸びた貧弱そのものだった可能性が高い。

(オフィアコドンの尾椎。筋肉の付く突起が弱い (#4))
これも語ると爬虫類と哺乳類のボディプランの比較から、両者の得手・不得手。それが生んだ適応放散の違いまで、とても記事一本では収まらないから、また次の機会にさせてほしい。楽しみにしていてくれ…
《胴体》

オフィアコドンの胴体は、一見すると面白味を感じられない。が、ヒトは見かけによらないとはよく言ったもので、探せば面白いことが見えてきた。
《基本情報》にて、オフィアコドンは水辺の魚食動物と筆者は記した。にも関わらず……
『オフィアコドンは泳ぎがド下手糞だった可能性があるのだ!!』
ファッ!?(# ゚Д゚)(# ゚Д゚)(# ゚Д゚)
(↑仰天する読者の図(筆者の予想))
そして読者は次にこう言う。
「いやいやいやいや!オカシイぞ!水辺の生物がカナヅチなはずがあるまい!!」
筆者も2年前に調べて仰天したのは良い思い出だ。たしかに指摘は正しい。クマやカワウソはもちろん、図体のデカいカバや普段は深みに入らないアオサギでさえ、必要と迫られれば見事な泳ぎを見せる。まだ信じられないようなので論文を紹介しよう。
2014年の研究(#2)、その名も『Was Ophiacodon (Synapsida, Eupelycosauria) a Swimmer? A Test Using Vertebral Dimensions (原題)/ オフィアコドンは遊泳家だったのか?(訳文)』
この研究では椎骨《Vertebral》を他の四肢動物と比較している(論文内ではメソサウルス《Mesosaurus》を始め、尻尾を推進力とする爬虫類にも触れられているため、“原初のウォータードラゴン”を知りたい方は読むと良いかもしれない)。以下は要約。
・古来よりオフィアコドンは半水生だったと考えられている(歯・四肢・組織学etcより)
・ただし、近年では上記の証拠の再解釈や再研究が進み、これは証拠の役を成さなくなってきた
・脊椎の中心の長さこそ半水生爬虫類と似ているが、他の最基盤の単弓類(100%陸生の種)もそれと似ていた。つまり、脊椎の特徴は決して半水生だった事を示しているわけではない
・その他の証拠も二次的な推測による曖昧なもののため、素直に陸生と考えるべきではないだろうか?
…半水生論者の心はメタメタのバッキバキに違いない。
傷口を抉るようで申し訳ないが、別の研究でもこれが間接的に支持されている。それは単弓類全体の椎骨の研究(#3)で、これによれば単弓類は皆、背骨を横に振る動きが苦手なことが示された。なるほど。たしかに私たちはヘビのようにクネクネさせる事は出来ない。程度の差はあれ、3億年前のオフィアコドンも同様だったとされている。
「だからどうした…」ではない。
これ、実は致命的である。何でも良いから哺乳類と鳥類以外の泳ぐ脊椎動物を思い浮かべてほしい。本当に何でも良いのだが、それは果たしてどのように泳いでいるだろうか?
……そうだ。察しの良い方もいるだろう。
メダカもマグロもイモリもヘビもワニも、みんな背骨を横に振って泳いでいるではないか。彼らには強い尻尾があり、それを左右に振ることで推進力を得ている。もちろんオフィアコドンにはペンギンのような前ビレ(フリッパー)はないし、クジラのように横に平たい尾ビレもない。だから当然推進力を得るには、身体を左右に振る必要があるのだ………が、肝心要の背骨が……動けん……馬鹿なッ!?
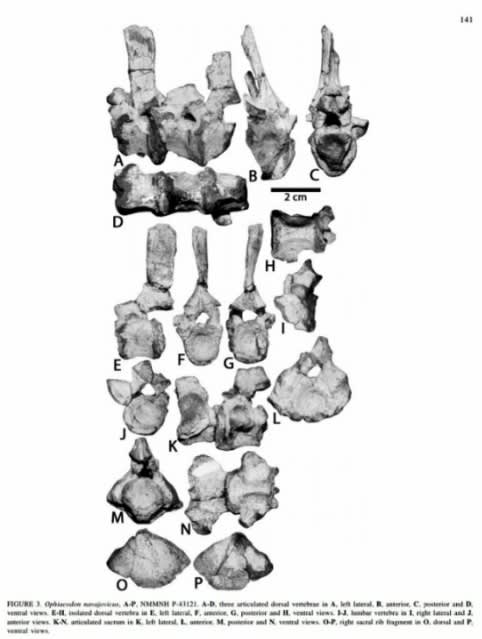
(オフィアコドンの椎骨など (#4))
…というわけだ。胴体の骨自体の特徴は、せいぜい上突起が背ビレ状になっておらず平坦であること。そして肋骨に括れが見られない(横隔膜は未発達)ことぐらいだろう。さして書くまでもないので、次に移りたい。
《四肢》
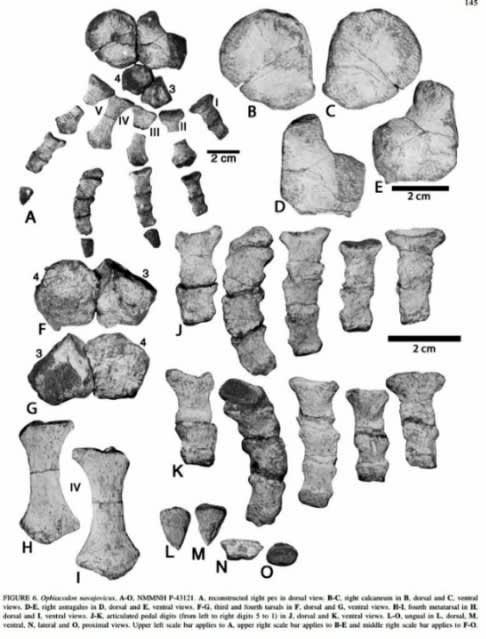
(オフィアコドンの手指など (#4))
オフィアコドンの四肢は短く、そして手指の骨が平たい。これは水辺で暮らしていたからだろう。持続的な推進力にはならないが、滑りやすい地面をしっかりと踏みしめたり、水底を蹴って滑るように動くことも出来たのではないだろうか。……というより、尻尾が貧弱なので、こうでもなければ水中で推進力できまい。そして、このような泳ぎ(?)では当然ながら速度は期待できないため、深みへ遠征したり、ワニ類よろしく獲物へ突撃するような真似は、残念ながら考えにくい。
…なんてアッサリ済むと思っていた筆者は馬鹿だった
なにせ他の大型単弓類(もちろんゴリゴリの陸生)も平たい手指の持ち主(#5)だったのだ。これはオフィアコドン以外の論文を読んで初めて知ったので、危うく皆さまへ誤解を与えるところだった(汗)
オイオイオイ(;・∀・)「どーゆーことやねん…」
ごもっともだ。どうやらかつては平たい手を根拠に水棲に特化していたとされていたらしい(←英Wikiより)が、ぶっちゃけ根拠と胸を張れるような代物ではない。筆者が思うに、こうした平たい手指は、初期の大型陸上四肢動物に共通していたのではなかろうか? 事実エリオプスからディメトロドンに至るまで、多くの種がこうした手指をしている(#6) こうした指は見るからに体重を支えやすそうだ。これらは論文はもとより各地の博物館でも確認できる。読者の皆さまも、コロナ禍が過ぎたらどうぞ見学してほしい。

(『大地のハンター展』より、エリオプスの胴体と四肢。指の太さと平べったさが分かるだろうか? (筆者撮影))
それと爪(末節骨)は鈍い。武器にはならなかっただろうし、何かをよじ登るのにも不適だった。身体の小さいアーケオティリス(←爪は見つかっていない)などは木登りが可能な鋭さもあったと思うが、少なくとも派生的な種類では上の写真の通りだ。ちなみに同時代のメセノサウルス類《mesenosaurine》(トカゲに似た有羊膜類)の場合、きちんと武器・登攀に使えそうな鉤爪が生えている。もしかすると、オフィアコドン類の平たい爪は、祖先から改めて進化し直した特徴だったのかもしれない。
では改め直して。
オフィアコドンを語る上で肩周り。とりわけ肩甲骨を外すことはできない。
(左側の平たい骨が肩甲骨 (#★))
なにせ頭骨を除けば、もっとも立派な骨なのだ。上部・下部・横幅ともに広い。莫大な量の筋肉が付着していた。なので生前は首周りが異様にムキムキだったと推測できる。そうした筋肉群は、ある物は大きな頭部を支え、ある物は四肢を動かす原動力となった。
やや想像を逞しくすれば、オフィアコドンが獲物を襲う際に前肢でスタートダッシュを切ったとも考えられるだろうか? あるいは獲物へ勢いよく頭を振り下ろしていたのだろうか?
ぶっちゃけ使い方までは分からない。ただ巷で囁かれるように、『哺乳類は前輪(脚)駆動・爬虫類は後輪(脚)駆動』という流れの原点を垣間見ることが出来る……という意味では重要な特徴だろう。
《頭部》

オフィアコドン(類)の頭部を見てまず最初に感じるのが、その大きさだろう。楕円形の頭骨は最大で50cm前後。高さも十分で、ずら〜っと並んだ歯も特徴的だ。まかり間違って噛みつかれるのは御免こうむりたい。
しかし幸いというべきか、オフィアコドン類に噛まれても大事には至らないだろう。なにせ彼らの歯は、先が尖っていても切れ味がない……つまり円錐形の歯をしていたのだ。オフィアコドン類は基本的に小動物か魚を狙うハンターであり、歯の第一の役割は獲物を滑り落とさないことだった(#8,#9)。このあたりは学名の由来(オフィアコドンとは“ヘビの歯”という意味)も示している。あいにく良質な資料は見つからなかったので、読者におかれては検索エンジンにて『teeth Ophiacodon』と検索してほしい。すると先述の円錐形をした歯を観察できる。

(『大地のハンター展』より、マレーガビアルの口先。縦横ともに最小限の抵抗となるようなデザインだ (筆者撮影))
こうした歯は、現生の魚や小動物を狙うワニ(↑のマレーガビアルなど)にも共通している。
オフィアコドン類の頭骨は左右幅が薄い。後頭部の咬筋の入るスペース(側頭窓)も狭かった。なので見た目ほど恐ろしい武器ではなかったようだ。とはいえ、非哺乳類型の単弓類は素早く噛む事が得意だったとする話(#8)もあり、おそらくオフィアコドン類の頭部へ求められたのは獲物を砕く力ではなく、素早く噛み付くスピードだったことを踏まえると、それも自然だろう……
……なんて終われればどんなに良かったことか。ところがどっこい、そうはいかないのがオフィアコドン。問屋もゲバ棒持って追い返すのが古生物学だ。
オフィアコドンの頭骨にもこれまた不可解な点が存在する。それは『頭骨の上下幅が高い』こと、そしてダメ押しに『鼻孔の位置が低い』ことだ。
冷静に考えてほしい。オフィアコドンの主食は間違いなく水辺の魚や両生類である。それらは何処に潜んでいるのかと言えば、もちろん水中だ。歯は問題ない。横幅も最小限に抑えられているから水の抵抗もない。
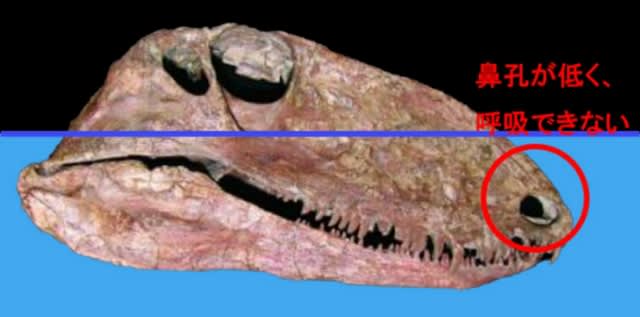
(オフィアコドンの頭部の問題点。水中では呼吸が出来ないし、振り回そうにも縦幅が邪魔だ (自作))
だがしかし!
ひとたび水中へ鼻先を突っ込んだら、もう悲惨だ。まず呼吸が出来ない。現生の半水生動物(カエル〜カバに至るまで)は、揃って鼻孔が水面へ突き出ている。いわば“動物版シュノーケル”だ。しかしオフィアコドンの鼻孔はその真逆。むしろ巻き上げた泥水を吸い込みかねない位置に開いている。もし無理やり鼻孔を外へ出そうとしたら、今度は身体全体が沈んでしまって話にならない ――そもそも頚椎の可動性からして難しいが。
なんとか呼吸の問題を解消したとしても、次は縦長の頭骨が待ち受ける。先ほどマレーガビアルの吩部を見せたのを覚えているだろうか? 水辺の捕食者の頭部は大別して2つ。1つはマレーガビアルやサギのような細長い“菜箸タイプ”か、あるいはカエルやワニのような平たい“カスタネットタイプ”だ。どちらも水中で獲物を捕える際に、水の抵抗を少しでも減らすための形態である。…にも関わらず、その逆を行った天の邪鬼がオフィアコドンその人。もう筆者も弁護のしようがない。なぜこんな不可解な頭部をしているのかは分からない。首は長くも無ければ特別柔軟でもないので、上空から勢いよく振り下ろす奇襲も難しかろう。あるいは、前述のガッシリした肩の筋肉群は、この矛盾に満ちた頭部を振り回すための“苦肉の策”だったのかもしれない……真相は不明だ。

(オフィアコドン類の1種ヴァラノサウルスの頭骨。口先の大きめな歯が犬歯である #)
やや余談に近いが、最初期のアーケオティリス《Archaeothyris》はもちろん、もう少し派生的なヴァラノサウルス《Varanosaurus》に至るまで、その多くが犬歯を長大化させているのは面白い特徴だ。形状は他の歯と変わらないのでほとんど誤差のようなものだっただろうが、ひょっとすると獲物を咥えた際にアンカーの役目を果たしたのかもしれない。とはいえ、最派生のオフィアコドン自体では犬歯が他の歯と同化して分かりにくくなっているので、元から大した意味はなかったのかもしれないが。
かくしてオフィアコドンの身体的特徴の解説は終わった。
あっ、わかったわかった! 皆さまの気持ちは十分伝わってますから、チョ待っア”ア”ア”ア”ア”ア”(流れ込む群衆
(# ゚Д゚)(# ゚Д゚)(# ゚Д゚)「こないな欠陥生物、さっさと滅んで当然やないかぁ!!」
うん。その通りだと首を縦に振りたいのは筆者も同じだ。しかし、また1つ朗報がある……
『オフィアコドン科は約3000万年も生存を続けていた』
これがどれだけ凄いかは、人類と比較してみると良い。人類の歴史は最古の猿人まで遡っても僅か700万年。オフィアコドンの歴史は、実にその4倍以上だ。まかり間違っても“欠陥生物”とは呼べまい。しかもオフィアコドンその間に、石炭紀の熱帯雨林の崩壊《Carboniferous rainforest collapse》(縮めてCRC)という絶滅事件をも生き延びていた。付け加えるなら、オフィアコドンは現状確認されている中では『最古の陸生大型肉食動物』でもある。
俄然面白くなってきた(某教授並み感)。
『何がオフィアコドン類を地球史上最初の陸棲大型肉食動物へ押し上げ、誰が彼らから初代王者の座を奪ったのか?』
言わば3億年に渡る『水際の興亡史』、その始まりを次回から探っていきたい。
次回『カナヅチワニもどき(後編)』にてお会いしよう。アディオーーース!
※白状すると、本日2021年7月15日に『水際の興亡史』が愛でたく発売となっている。後追いだの便乗だのは願い下げだったので、急ぎで書き上げた次第だ。ところどころボロが目立つが、まぁ……笑ってほしい。あるいは発売記念ということで1つw
《余談》
オフィアコドンの代謝について怪しくも面白い話がある。2017年の長骨(※四肢の骨)の論文(#11)によれば、彼らの代謝は原始的な外見とは裏腹に内温・温血動物のそれと近いもので、成長スピードも速かったらしい。にわかには信じがたいのだが、筆者の中で一つ、しっくりくる説明がある。
皮革業者の間では、昔から効率よくワニ革(もちろんワニの代謝は低い)を手に入れるためにある工夫をしている。それは飼育場の気温をガンガンに上げ、餌も山盛り与えることだ。こうすると変温動物でも代謝が促進され、成長も速まる。
これと同じことが当時の地球環境に起こったのかもしれない。詳しくは次回取り上げるが、当時の水辺はオフィアコドンにとって楽園だった。餌は多ければ天敵も少なく、気温も安定していた。まさに楽園であり、それがためにオフィアコドンは急速な成長と高い代謝の維持が可能だったのかもしれない……信じるか信じないかは、アナタしだい(・∀・)9
《参考文献》
[論文]
#1『A multitaxic bonebed near the Carboniferous–Permian boundary (Halgaito Formation, Cutler Group) in Valley of the Gods, Utah, USA: vertebrate paleontology and taphonomy』(Adam K Huttenlocker:2018)…石炭紀〜ペルム紀の生物相
#2『Was Ophiacodon (Synapsida, Eupelycosauria) a Swimmer? A Test Using Vertebral Dimensions』(Ryan N Felice:2014)…オフィアコドンの椎骨
#3『Adaptive landscapes challenge the “lateral-to-sagittal” paradigm for mammalian vertebral evolution』( Katrina E. Jones:2021)…単弓類の椎骨の可動性の進化
#4『A PARTIAL SKELETON OF OPHIACODON NAVAJOVICUS (EUPELYCOSAURIA: OPHIACODONTIDAE) FROM THE UPPER PENNSYLVANIAN OF CAÑON DEL COBRE, NEW MEXICO』(Susan Harris:2010)…オフィアコドンの下半身
#5『Patterns of evolution in the manus and pes of non-mammalian therapsids』(James A. Hopson:2010)…単弓類の手指の進化
#6『The behavioral and biostratigraphical significance and origin of vertebrate trackways from the Permian of Scotland』(Patrick J McKeever:1994)…スコットランド産の四肢動物の足跡化石
#7『Permo—Carboniferous Paleoecology and Morphotypic Series』(Everett C Olson:1975)…単弓類の生息地と頭骨の変遷
#8『Evolutionary Patterns in the History of Permo-Triassic and Cenozoic Synapsid Predators』(Blaire Van Valkenburgh:2002)…肉食性単弓類の進化
#9『Microanatomy of the radius and lifestyle in amniotes (Vertebrata, Tetrapoda)』(Damien Germain:2005)…頭骨から探る有羊膜類の生態
#10『A reevaluation of early amniote phylogeny』(MICHEL LAURIN:1995)…有羊膜類の頭部
#11『Ophiacodon long bone histology: the earliest occurrence of FLB in the mammalian stem lineage』(Christen Shelton:2015)…オフィアコドンの代謝
#12『Limb-Bone development of seymouriamorphs: implications for the evolution of growth strategy in stem amniotes』(Jordi Estefa:2020)…初期の四肢動物の成長速度
#12『Dimetrodon Is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution』(Kenneth D Angielczyk:2009)…単弓類の総括)
#2
WEB
#★『Ophiacodon uniformis (Cope, 1878)』…オフィアコドンの基本情報と骨格
[洋書]
#A『Ophiacodon (Synapsida, Ophiacodontidae) from the Lower Permian Sangre de Cristo Formation of New Mexico AMYC 』(DAVIDS BERMAN:2013)
#B『A PARTIAL SKELETON OF OPHIACODON NAVAJOVICUS (EUPELYCOSAURIA: OPHIACODONTIDAE) FROM THE UPPER PENNSYLVANIAN OF CAÑON DELCOBRE, NEWMEXICO 』(SUSANK HARRIS:2010)
#C『RE-EVALUATION OF RUTHIROMIAELCOBRIENSIS (EUPELYCOSAURIA: OPHIACODONTIDAE?) FROM THE LOWER PERMIAN (SEYMOURAN?) OF CAÑONDELCOBRE, NORTHERN NEWMEXICO』 (JUSTINA SPIELMANN:2010)
#D『Forerunners of Mammals: Radiation • Histology • Biology』(Chinsamy-Turan:2011)
[和書]
・『哺乳類型爬虫類-ヒトの知られざる祖先』(金子隆一:1998)
・『絶滅哺乳類図鑑』(富田幸光:2002)
・『恐竜異説』(ロバート・バッカー:1989)
・『肉食恐竜事典』…(グレゴリー・ポール:1993)
・『生命大躍進』(図録)…(科博:2015)