









スピノやダケント復元教室のレポートは書かず、かといって「墓場」や「ハツェグ島」の続きは渋っている状況。そんなワケで今回は、筆者の気まぐれにより、地球最古の陸棲巨大生物を取り上げようと思う。
「背ビレ竜、ディメトロドンかい?」
ナンセンスだ。
「ゾワゾワの王、アースロプレウラ?」
それも違う…けど嫌いじゃない。
…そんな物理的に低い連中ではない。
今回取り上げるのは、獣弓類の先駆けディノケファルス類《Dinocephalia》である!!

(↑ディノケファルス類の1種、エステメノスクスの組み立て骨格 ©NHK)
ダイレクトに和名を恐頭類(「ディノ=恐るべき」×「ケファルス=頭」)と書き、それは言わずもがな一目で納得するだろう。多くの種が太く長い牙を持っていながら、それすら見劣りさせるほどの装飾(ヘルメット状の瘤や扇状の突起)で頭を飾り立てている。オオツノジカなどの規格外な偶蹄類を別とすれば、史上最も派手な“単弓類”と呼んで差し支えあるまい。
ちなみに、ここで出てきた“単弓類”と呼ばれる分類群について補足すると、彼らは私たち哺乳類の祖先(より正確には哺乳類を内包する分類群)で、ちょうどニワトリから見たティラノサウルスをイメージすると分かりやすいだろう(※分類的な立ち位置の話)。

(↑大地をのし歩くディノケファルス類 ©NHK)
ただ、筆者が書き連ねたいこと(生態etc)とは別に、どうしても書かずにはいられなかった情報がある。…何を隠そう、ディノケファルス類の「分類」についてだ。
現行の日本語版Wikipediaや当ブログの単弓類において、数少ない日本語の参考文献となっている(著)金子隆一の『哺乳類型爬虫類 -ヒトの知られざる祖先』では、ディノケファルス類を異歯亜目(ディキノドン類etc)の仲間としている。ところが、1997年の論文(#2)では両者に直接の関係はないとされている。正直なところ分類云々は筆者もさっぱり ――私見では異歯亜目よりかは、むしろ獣弓類全体の最基盤っぽい―― であるし、今回の大筋には関わってこないため、これ以上は深入りしないことにする。
興味のある方は、Googlescholarで「Anomodontia(異歯亜目)」とでも打ち込んで調べてくださいお願いしますワタクシガシンデシマウノデ(殴
閑話休題。そんなディノケファルス類には、大別して2つの系統が存在する。1つは禍々しい顎と牙を備えたアンテオサウルス上科《Anteosauroidea》で、もう1つは横綱も真っ青な体格のタピノケファルス科《Tapinocephalia》だ。
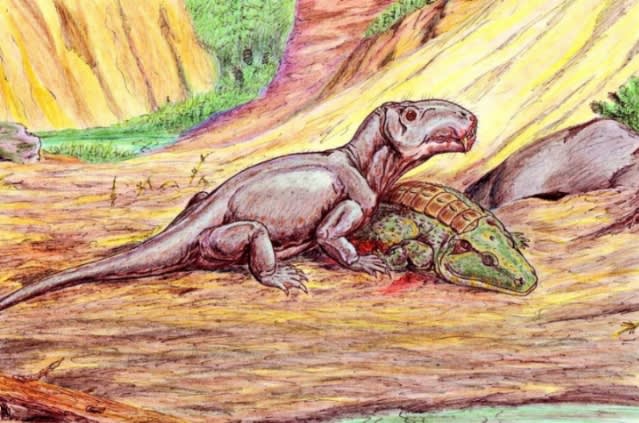
(↑両生類を捕らえたアンテオサウルス科のブリソプス《Brithopus》 Wikiコモンズより)

(↑茂みの中のタピノケファルス科 ©NHK)
多少の議論はあるものの、概ねアンテオサウルス科が肉食で、タピノケファルス科が植物食とされている(近日投稿予定の補記を参照されたし)。どちらも鎧兜のような頭と重心安定した胴体を持っていた。対称的に尻尾は貧弱でものの役にも立ちそうにない。これだけ聞くとカバっぽく思えるが、その四肢は斜め横に張り出しており、現存の生物に類似の種は存在しないだろう。
ここまで特異な姿をしたディノケファルス類には、かねてより1つの“定説”というか、旧時代的な解釈が行われてきた。それは…
『ディノケファルス類がカバのような半水棲だったのではないか…?』
なんて疑惑だ。
筆者なりの結論を言えば、こちらは『否!!』(# ゚Д゚)……である。
より正確には、ディノケファルス類は水辺を好んだものの、年がら年中プカプカ半身浴をしてはいなかった…と言おう。
このあたりの話を掘り下げると、それはそれで深くなってしまう ――下手すりゃ記事1〜2本分になる―― ため、ここでは半水棲説を否定する有力な証拠を1つ挙げるに留めたい。その証拠とは、ズバリ眼窩と鼻孔の位置だ。

(↑モスコプスの頭骨。眼窩は見事に水平に開いている Wikiコモンズより)
見ての通り、ディノケファルス類の眼窩は真横を向いており、鼻孔は鼻先の中腹に付いている。では読者の皆さまに質問しよう。この状態で水に浸かったらどうなってしまうのか……? 賢明なる紳士淑女であれば悩むまでもない。

(↑水面に顔を出したカバ。水中で快適に過ごすには、このような突出した目が必要なのだ。 提供:@StarPredator247氏)
ということで、少なくとも今回に限っては、ディノケファルス類が陸をメインに活動する動物だとして話を進めていかせてもらう。
ディノケファルス類の黄金時代は、今から約2億6千5百万年前のペルム紀中期〜後期(バイオゾーン上のタピノケファルス帯《Tapinocephalus Assemblage Zone》)とされている。この時代、陸地は超大陸パンゲアによってのみ構成されていた。大陸の内陸部で砂漠化が進行する一方、沿岸部には石炭紀から引き続いて安定した降水が提供され続けており、これが初期の裸子植物から、ひいては生態系の頂点に立つディノケファルス類を支えていたのである。

(↑ペルム紀中期〜後期の世界。大陸の沿岸部には肥沃な土地が拡がっていた ©NHK)
当時の陸上では非常に興味深い現象が発生していた。なんと(地質学的なスケールにおいて)昨日まで繁栄を極めていた初期の単弓類(俗に言う盤竜類)や陸棲特化の両生類が次々に姿を消し、生態系の上位に空白が目立つようになっていたのだ。この謎めいた事件はオルソンの空白《Olson's Extinction》と呼ばれていて、今も原因が探られている(#4)。
そんな千載一遇の機に乗じ、我らが主役ディノケファルス類は多様化を成し遂げた。己が巨体を頼みに、抵抗勢力をニッチという名の土俵から寄り切り、世界を半ば独占したのである。川原で寝転んでいるのはディノケファルス類、その奥で腐肉を漁っているのもディノケファルス類、ちょうど森から顔を覗かせたのもディノケファルス類……なんてことがザラにある世界。それが2億6千5百万年前の日常だった。
こうした大繁栄のさなか、その立役者ディノケファルス類は、ある革新的な進化を遂げていた。だが不思議なことに(翻訳書を含めて)、それを和書で取り上げられたところは齢十八を数える筆者でさえ見たことがない。
それは単純な巨大化ではない。たしかに最大級のディノケファルス類は体重850kgに達したとする研究(#3)もあって、これは陸上生命史における快挙だ。
しかし、さらに重要なのが背丈が高くなったことなのだ。
それまでのエダフォサウルスやディアデクテスのような植物食動物は、揃いも揃って低身長 ――遠目にはオオトカゲにしか見えない―― の種類ばかりであった。当然だろう。なぜなら彼らは、見たまんま90°のガニ股を貫く姿をしており、短足も相まって物理的に低い生物だったのだ。手が届いたとして、せいぜい地上から4〜50センチもあるまい。
とはいえエダフォサウルスなどの台頭は、植物にとって厄災以外のなんでもなかった。菜食主義の四足動物が増えていくにつれ、植物全体の多様性は低くなっていたことがニール・ブロックレハースト(Neil Brocklehurst)らの研究(#5)により、今年になって明らかにされた。
それでも、彼らの影響は丈の低いシダ類などに限られていた。数メートルの超える初期の樹木は、未だに安寧と日光を享受していたのである。

(↑モスコプスの組み立て骨格 Wikiコモンズより)
ところがディノケファルス類に関しては、もはやオオトカゲなどと笑っていられない。際立った背丈は、大台1mを一気に飛び越え、最大級の種類では2m近い高さにさえ届くようになっていた。当然メートル級のトクサだのロボクだのにも、あっさり食指が伸びただろう。
何度も繰り返すようで申し訳ないが、ディノケファルス類の歯と顎は前時代の生物の比にならないほど強力無比だった。しかもタピノケファルス科の歯列は、後の鳥盤類や有蹄類の歯列を先取りした使い勝手の良い形状であったことが、昨年の研究(#6)で明らかとなっている。
たとえ繊維質の多いトクサだろうが、鎧のようなロボクの樹皮だろうと、なんであれ紙切れ同然に噛み千切ったはずだ。

(↑古生代のトクサ類《Annularia》の復元図 Wikiコモンズより)
あいにくディノケファルス類の咬合力(顎を閉じる力)を具体的に数値化した研究は見つからなかった ――もし貴方が知っているなら、是非ともコメントで教えていただきたい(´・ω・`)。
だが砲弾の如き頭部や補強された口蓋を見れば、どんなに控えめに見積もろうと、ツキノワグマに匹敵する咬合力(200kg)を生み出せたであろうことは、想像に難くないだろう。
つまるところ決論としては、ディノケファルス類の台頭によって樹木にも本格的な淘汰圧が促された可能性があると、筆者は考えている((#7)の論文も参照されたし)。
ディノケファルス類によって伐採された土地は、シダを始めとした下草にとって理想的な生育地となっただろうし、エオディキノドン《Eodicynodon》など初期の穴居性動物も巣穴を構えやすかろう。この辺は、近いうちに南半球の研究者(とりわけアフリカ)の頑張りに期待したいところだ。いつの日にか、上述の内容が論文化されることを祈ってやまない。
ではここで話を180°切り替え、読者の皆さまへ再び質問といこう。ディノケファルス類の得意技をご存知だろうか? ヒントはゴツゴツした頭部である。
…御名答!! まさしく頭突きだ。こちらは百聞は一見にしかずなので、ひとまず実骨を観察してほしい。

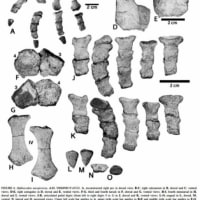
(↑A,アンテオサウルス、B,タピノケファルス科のウレモサウルス《Ulemosaurus》の頭骨。 コブ状の部位が確認できる #1より)
彼らの頭部に対する学者の意見(#8)は、研究黎明期からほとんど変わっていない。グダグダ議論までもなく、火を見るより明らかであろう。もしディスプレイ用の虚仮威しだとしたら、それはエネルギーの浪費と言わざるをえない。なお瘤の骨密度は高かったため、ぶつけ合うに足る強度も十分に備えていた。
こうした“恐ろしい(ディノ)頭(ケファルス)”は、過去〜現代にかける様々な生物にも見られる。過去ならば“石頭恐竜”パキケファロサウルス類、現代ならば極圏のジャコウウシや高山のオオツノヒツジ(ビッグホーン)といった具合いに。

(↑パキケファロサウルスの頭骨)

(↑ジャコウウシの決闘)
どれも程度の差はあれど、ヘルメット状の顕著に盛り上がった頭頂部(と棘や角)をしている。
ちなみにディノケファルス類こそ直接的に触れられていないものの、こうしたドーム状の頭部を持つ動物には、何かしらの収斂進化が働いていたのではないか? …とする研究(#9)もあり、非常に興味深い。
ただし、前述の堅頭竜類や偶蹄類とディノケファルス類には、決定的に異なる点があった。これも目敏い読者ならば気づくだろう。それは彼らの四脚である。

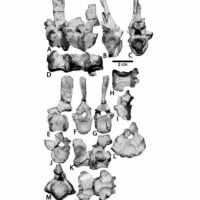
(↑ディノケファルス類の四脚 Wikiコモンズより)
ディノケファルス類の脚を見て気づく事といえば、その短足さ加減だろう。この四肢では回転数を上げてドタタタタッ!と走ることはできても、決してタタッ!タタッ!タタッ!っと、軽快なステップを刻むことは望めそうもない。しかも肘や膝はガニ股〜中腰で固定されているため、ジャコウウシよろしく、真っ直ぐ走って相手の頭頂部ピッタへ激突するのも難しい。
ディノケファルス類の名誉のために言っておくと、彼らの四脚は、一昔前の学者に鼻で笑われていたような貧弱な脚ではない。ゴツゴツした上腕骨や大腿骨が、何よりの証である(言うまでもなく筋肉質だったことの証だ)。
…しかしだ。彼らの脚がダッシュアタック向きではないことは、火を見るより明らかであり、『哺乳類型爬虫類-ヒトの知られざる祖先』でも同様のことが指摘されている。これがもし、ジャコウウシやオオツノヒツジのようにダッシュアタックを得意技とする生物であれば、あまりにも不自然極まりない。
となると、ディノケファルス類の決闘はジャコウウシなどのそれとは趣きの異なるものだったに違いない。
ディノケファルス類を正面から眺めると、横に張り出した四肢のおかげで、おおよそ頭を頂点とした三角形ないし台形に見えるだろう。この体型は重心を安定させやすく、立派な肥満体型を支えたり、頭突き相撲において踏ん張ったりするにも都合が良かったはずだ。それこそ四股を踏んだ力士に近い、抜群の安定性を誇ったことだろう。

(↑頭突きを行うタピノケファルス類 ©NHK)
現生のウミイグアナも、オス同士が頭突きを行うことが知られている。こちらも行儀よく向かい合ってから、ガニ股で踏ん張りつつ、ジワジワと押し合う方法だ。やはり間違ってもジャコウウシのようなダッシュアタックなどしない。
ただし筆者は、ディノケファルス類の決闘が頭突きによるもののみだったとは、正直疑わしいと思っている。では他に何を武器にしてたかというと、ディノケファルス類をディノケファルス類足らしめている特徴……そう、発達した牙だ。
なにも突拍子も無い話ではない。一部のディノケファルス類(とりわけアンテオサウルス科)の牙は、前方へ向かって飛び出していた。ちょうど明石家さんまをイメージすると良いかもしれない。
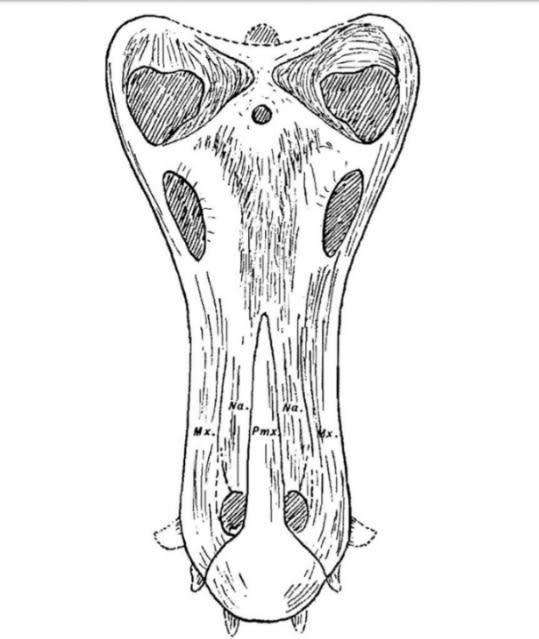
(↑ジョンケリアの頭骨スケッチ。数本の前歯が突出している #10より)
現生のカバの前歯がそうであるように、ディノケファルス類もまた、この出っ歯で仲間同士ド突きあったりしていたと考えても、なんら不思議ではない。現にカバは同様の歯を用いて壮絶な噛み合いを繰り広げている(詳しくは資料映像を閲覧されたし)。

(↑牙を突き立てながら争うカバ)
当然ながら、そんな歯(と強靭な顎)で噛まれては無事で済むはずがない。実際ディノケファルス類の1種ジョンケリア《Jonkeria》の大腿骨には、外敵(おそらく他のディノケファルス類)の噛み傷が刻まれていた。それを報告した論文(#11)中では、傷を残したのは捕食者であるとされている。だが大概のディノケファルス類の備えた破壊的な牙を踏まえると、どうしても仲間内における諍いの可能性を筆者は捨てきれない ――にしては傷が深いし、噛まれた箇所も不自然だが。

(↑捕食者の餌食となったディノケファルス類 ©NHK)
このように、陸上生態系において、初めて活動的(恒温的とも言えよう)な世界を創り上げたディノケファルス類であったが、皮肉にも在りし日の自らが為した簒奪の因果によってか、彼らもまた簒奪の憂き目に遭うことになった。
ディノケファルス類がタピノケファルス帯に黄金時代を築くのと、ほぼ時を同じくして、やはり進化の荒波に揉まれた ――それを強いたのは王者ディノケファルス類であった―― 肉食獣の獣歯類《Theriodontia》(ゴルゴノプス亜目&テロケファルス亜目)や草食獣の異歯亜目《Anomodontia》(ディキノドン類etc)が本格的な多様化を始めている。これら新鋭の獣弓類は、どれも強靭な顎と柔軟な身体を備え、ディノケファルス類よりも敏捷で適応放散に長けていた。
たしかにディノケファルス類も各ニッチにおいて一定の繁栄を見せたものの、その外見は良くも悪くも似たり寄ったりで、棲み分けという意味では十分な成功を収められずにいたのだろう。
かつてロバートバッカーが『恐竜異説』において述べたように、ひとたび解き放たれた進化の悪魔は、生物のアグレッシブ化によって歯止めを失い、その根源たるディノケファルス類さえも容赦なく淘汰されていったのである。2018年の研究(#12)でも、時代が下るにつれて生物種の絶滅率が高くなっていたことが示されており、それだけ生存競争が激化していたようだ。
こうした新勢力の台頭に合わせるようにディノケファルス類が衰退していく様は、まるで巨大なれど旧時代的な戦艦が、小型で小回りの効く戦闘機に敗れたのを彷彿とさせてくれる。

(↑獲物を咥えたゴルゴノプス亜目のリカエノプスと、川原を行くディキノドン類)
かくしてタピノケファルス帯の末期をもち、ディノケファルス類の血筋は完全に断絶した。
だがそれでもなお、彼らの築いた陸上生態系の基礎は、他の獣弓類→クルロタルシ類→恐竜→現生哺乳類へ、脈々と受け継がれていき、世界を仁義なき修羅地獄へと導いていく(#13)。
…この星を初めて“のし歩いた”ディノケファルス類。彼らの再評価が行われる日は、そう遠くないのかもしれない。
《資料映像》
※余談1
話の都合で誤解が生まれてしまいそうなので忠告しておく。鈍重そうな見た目に騙されてはいけない!! カバは小突くだけでナイルワニを蹴散らし、ライオンの群れ相手に千切っては投げを続け、果ては時速40kmで自動車を猛追する生きた戦車なのだ。さすが“アフリカで最も危険な野獣”である。
※余談2
Wikipediaなどを覗いていると(ディノケファルス類に限らないのだが)、胸部の組み立てが失敗している骨格を見つけることが多い。ティタノフォネウス(英語版)のサムネなど、まさに代表例で、あれでは体重を支えることが出来ない。今回の記事立てにあたり使用した組み立て骨格は、なるべく違和感のない骨格を使用としたつもりだが、念のため読者個人も胸部の作りを見当し直してほしい。
《資料映像》
《参考文献》
[論文]
#1『Dimetrodon Is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution(Kenneth D Angielczyk:2009)』(単弓類の総括)
#2『Dinocephalians are not anomodonts(FE Grine:1997)』(分類云々)
#3『First 3D reconstruction and volumetric body mass estimate of the tapinocephalid dinocephalian Tapinocaninus pamelae (Synapsida: Therapsida)(Marco Romano:2019)』(タピノケファルスの体重)
#4『Olson's Gap or Olson's Extinction? A Bayesian tip-dating approach to resolving stratigraphic uncertainty(Neil Brocklehurst:2020)』(オルソンの空白)
#5『The origin of tetrapod herbivory: effects on local plant diversity(Neil Brocklehurst:2020)』(植物の淘汰)
#6『Histological and developmental insights into the herbivorous dentition of tapinocephalid therapsids(Christian A Sidor:2019)』(タピノケファルス科の歯列)
#7『Functions of phytoliths in vascular plants: an evolutionary perspective
(Caroline AE Strömberg:2016)』(植物の進化)
#8『Synchrotron scanning reveals the palaeoneurology of the head-butting Moschops capensis (Therapsida, Dinocephalia)(Julien Benoit:2017)』(頭突き説)
#9『A Dome-Headed Stem Archosaur Exemplifies Convergence among Dinosaurs and Their Distant Relatives (Michelle R. Stocker:2016)』(頭突きの収斂進化)
#10『On the carnivorous mammal-like reptiles of the family Titanosuchidae(R Broom:1929)』(ジョンケリアの半身)
#11『Osteomyelitis in a 265-million-year-old titanosuchid (Dinocephalia, Therapsida)(Christen D Shelton:2017)』(ジョンケリアの負傷)
#12『Evolutionary rates of mid-Permian tetrapods from South Africa and the role of temporal resolution in turnover reconstructionMICHAEL O. DAY ET AL. MID-PERMIAN TETRAPOD …(Michael O Day:2018)』(四足動物の進化と絶滅)
#13『Evolution of the Permian and Triassic tetrapod communities of Eastern Europe(AG Sennikov:1996) https://t.co/4MTnRQToOJ』(ペルム紀〜三畳紀の食物網)
[和書]
・『哺乳類型爬虫類-ヒトの知られざる祖先』
・『恐竜異説』
・『絶滅哺乳類図鑑』




※コメント投稿者のブログIDはブログ作成者のみに通知されます