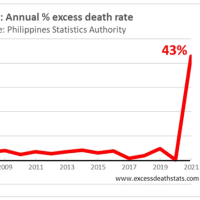
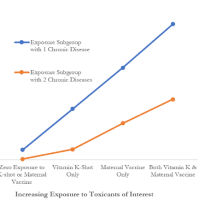
発熱は、免疫を強化し、多くの感染症の生存率と回復の改善に関連しているという証拠
解熱剤を使用してはいけない理由、そしてなぜ「放っておく」べきなのか
の文献4, 5, 6, 7, 8, 20, 25 Nat Rev Immunol. 2015 Jun; 15(6): 335–349
の要旨と冒頭部分の翻訳
+++++++++++++++++++++++++++
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4786079/
Nat Rev Immunol. 2015 Jun; 15(6): 335–349.
Published online 2015 May 15.
doi: 10.1038/nri3843
PMCID: PMC4786079
NIHMSID: NIHMS755275
PMID: 25976513
Fever and the thermal regulation of immunity: the immune system feels the heat
発熱と免疫の温度調節:免疫系は熱を感じます
Sharon S. Evans, Elizabeth A. Repasky, and Daniel T. Fisher
The publisher's final edited version of this article is available at Nat Rev Immunol
Abstract要旨
Fever is a cardinal response to infection that has been conserved in warm and cold-blooded vertebrates for over 600 million years of evolution.
発熱は感染症に対する基本的な反応であり、6 億年以上の進化の間、温血脊椎動物と冷血脊椎動物に保存されてきました。
The fever response is executed by integrated physiological and neuronal circuitry and confers a survival benefit during infection.
発熱反応は統合された生理学的回路と神経回路によって実行され、感染時の生存に利益をもたらします。
Here, we review our current understanding of how the inflammatory cues delivered by the thermal element of fever stimulate innate and adaptive immune responses.
ここでは、発熱という熱要素によってもたらされる炎症の合図がどのように自然免疫応答と適応免疫応答を刺激するかについて、現在の理解をレビューします。
We further highlight the unexpected multiplicity of roles of the pyrogenic cytokine interleukin-6 (IL-6), both during fever induction as well as during the mobilization of lymphocytes to the lymphoid organs that are the staging ground for immune defence.
さらに、発熱誘発性サイトカインであるインターロイキン 6 (IL-6) の役割が、発熱誘発時と免疫防御の拠点であるリンパ器官へのリンパ球動員時の両方で、予想外に多様であることを強調します。
Finally, we discuss the emerging evidence that suggests the adrenergic signalling pathways associated with thermogenesis shape immune cell function.
最後に、熱産生と免疫細胞の機能に関連するアドレナリン作動性シグナル伝達経路を示唆する新たな証拠について説明します。
++++++++++++++++++++++++
The fever response is a hallmark of infection and inflammatory disease and has been shaped through hundreds of millions of years of natural selection.
発熱反応は感染症や炎症性疾患の特徴であり、何億年にもわたる自然選択によって形作られてきました。
Febrile temperatures are so closely linked to the inflammatory response that heat (calor) is one of the four cardinal signs of inflammation, along with pain (dolor), redness (rubor), and swelling (tumour), as described by Celsus in ~30 BC.1
発熱は炎症反応と非常に密接に関連しているため、熱(熱)は、痛み(痛み)、発赤(発疹)、腫れ(腫瘍)と並んで、炎症の 4 つの主要な兆候の 1 つであると、ケルススは約 BC30 年に説明しています。1
The induction of fever in endothermic (warm-blooded) animals occurs at a high metabolic cost such that a 1°C rise in body temperature requires a 10–12.5% increase in metabolic rate.2
内温動物(温血動物)の発熱の誘発には高い代謝コストがかかり、体温が 1℃上昇すると代謝率が 10 ~ 12.5% 増加する必要があります。2
There is mounting evidence that the increase of 1 to 4°C in core body temperature that occurs during fever is associated with improved survival and resolution of many infections.
発熱中に起こる中核体温の 1 ~ 4°C の上昇が、多くの感染症の生存率と回復の改善に関連しているという証拠が増えています。
For example, the use of antipyretic drugs to diminish fever correlates with a 5% increase in mortality in human populations infected with influenza virus and negatively affects patient outcomes in the intensive care unit.3–5
たとえば、熱を下げるために解熱剤を使用すると、インフルエンザウイルスに感染したヒト集団の死亡率が 5% 増加することと相関しており、集中治療室での患者の転帰に悪影響を及ぼします。3-5
Preclinical studies in rabbits infected with rinderpest virus also found an increase in mortality when fever was inhibited with the antipyretic drug acetylsalicylic acid — 70% of acetylsalicylic acid-treated animals died as a result of infection as compared with only 16% of animals with a normal febrile response.6
牛疫ウイルスに感染したウサギを対象とした前臨床研究でも、解熱薬アセチルサリチル酸で発熱を抑制すると死亡率が増加することが判明した
アセチルサリチル酸で治療した動物の 70% が感染により死亡しましたが、正常な発熱反応を示した動物はわずか 16% でした。6
However, fever is not universally beneficial, particularly in cases of extreme inflammation where lowering, rather than raising body temperature has evolved as a protective mechanism.7–10
しかし、発熱は普遍的に有益というわけではなく、特に極度の炎症の場合には、体温を上げるのではなく体温を下げることが防御機構として進化している7-10。
Thus, uncontrolled fever is associated with worse outcomes in patients with sepsis or neurological injuries, whereas treatments that induce hypothermia can have a clinical benefit.11,12
したがって、敗血症または神経損傷を患う患者では、発熱が制御されていないと転帰が悪化しますが、低体温を誘発する治療には臨床上の利点がある可能性があります。11,12
A challenge in ascertaining the precise value of fever in endotherms is that the antipyretics used to inhibit fever target multiple aspects of the inflammatory response besides temperature regulation.11
内温における発熱の正確な値を確認する際の課題は、発熱を抑制するために使用される解熱剤が、体温調節以外にも炎症反応の複数の側面を標的にしていることです。11
Ectothermic (cold-blooded) vertebrates, which last shared a common ancestor with mammals over 600 million years ago, provide an ‘experiment in nature’ in which to examine the direct impact of febrile temperatures on survival.
変温動物(冷血)脊椎動物は、6億年以上前に最後に哺乳類と共通の祖先を共有しており、生存に対する発熱性の直接的な影響を調べるための「自然界での実験」を提供します。
Ectotherms as diverse as reptiles, fish, and insects raise their core temperature during infection through behavioural regulation, which leads to their seeking warmer environments (despite the risk of predation) or, in the case of bees, raising the local temperature of the hive through increased physical activity.2,13–19
爬虫類、魚類、昆虫などの多様な外温動物は、感染中に行動制御を通じて中核体温を上昇させ、その結果、(捕食の危険にもかかわらず)より暖かい環境を求めることになります。
ミツバチの場合、身体活動の増加により巣の局所温度が上昇します。2、13–19
Landmark studies published 40 years ago by Kluger’s laboratory showed that survival of the desert iguana Dipsosaurus dorsalis is reduced by 75% if prevented from behaviourally raising its core temperature by approximately 2°C after infection with the Gram-negative bacterium Aeromonas hydrophila.2,13,14
クルーガーの研究室が 40 年前に発表した画期的な研究では、グラム陰性菌エロモナス ハイドロフィラに感染した後、行動的に中核体温が約 2℃上昇するのを妨げると、サバクトビグアナ ディプソサウルス ドーサリスの生存率が 75% 減少することが示されました。2,13 ,14
The heat-seeking behaviour of the desert iguana, blue-finned tuna and leech is negated by antipyretic drugs, indicating that common biochemical pathways drive fevers in ectothermic and endothermic animals.14,16,20
サバクイグアナ、クロマグロ、ヒルの熱を求める行動は解熱薬によって打ち消され、共通の生化学的経路が変温動物と内温動物の発熱を引き起こすことを示しています。14、16、20
Surprisingly, the correlation between infection and increased temperature extends even to plants, which arose 1.5 billion years ago.
驚くべきことに、感染と気温上昇との相関関係は、15億年前に誕生した植物にまで及びます。
For example, the temperature of the leaves from the bean plant Phaseolus vulgaris increases by around 2°C following infection with the fungus Collectotrichum lindemuthianum.21
たとえば、マメ科植物インゲンマメの葉の温度は、真菌 Collectotrichum lindemuthianum に感染すると約 2°C 上昇します。21
Thermoregulation in plants occurs through mitochondrial respiration22, although it is not known whether these fever-like responses have a direct impact on clearance of infection.
植物の体温調節はミトコンドリア呼吸によって行われます 22 が、これらの発熱のような反応が感染の排除に直接影響を与えるかどうかは不明です。
The fact that fever has been retained throughout vertebrate evolution strongly argues that febrile temperatures confer a survival advantage.
脊椎動物の進化を通じて発熱が維持されてきたという事実は、高熱が生存に有利であることを強く主張しています。
A long-standing mystery relates to the protective mechanisms by which fever wards off attacks by invading pathogens.
長年の謎は、侵入する病原体による攻撃を発熱が防ぐ防御機構に関係している。
One mechanism involves direct effects of febrile temperatures on the infectious potential of pathogens.23
メカニズムの 1 つは、病原体の感染力に対する高温の直接的な影響です。23
For example, temperatures in the febrile range (40–41°C) cause a greater than 200-fold reduction in the replication rate of poliovirus in mammalian cells and increase the susceptibility of Gram-negative bacteria to serum-induced lysis.24,25
たとえば、発熱範囲(40 ~ 41°C)の温度では、哺乳動物細胞におけるポリオウイルスの複製速度が 200 分の 1 を超えて低下し、血清誘導性溶解に対するグラム陰性細菌の感受性が増加します。24、25
In this Review, we discuss the evidence suggesting that febrile temperatures boost the effectiveness of the immune response during infections by stimulating both the innate and adaptive arms of the immune system.
この総説では、高温が免疫系の先天的および適応的部門の両方を刺激することにより、感染時の免疫反応の有効性を高めることを示唆する証拠について議論します。
We will highlight the role of the pyrogenic cytokine interleukin-6 (IL-6) in two key phases of the febrile response: firstly in driving the rise in core temperature, and secondly as a downstream effector cytokine orchestrating lymphocyte trafficking to lymphoid organs.
我々は、発熱反応の 2 つの重要な段階における発熱性サイトカイン インターロイキン 6 (IL-6) の役割に焦点を当てます。1 つは深部体温の上昇を促進する役割、もう 1 つはリンパ器官へのリンパ球輸送を調整する下流のエフェクター サイトカインとしての役割です。
We also describe febrile temperature as a ‘rheostat’, dialling down systemic inflammation during the return to homeostasis.
また、発熱性の体温は、恒常性への復帰中に全身の炎症を抑える「レオスタット」とも表現されます。
Finally, we highlight new data demonstrating the overlapping signalling pathways that are involved in thermogenesis and in the regulation of the immune response.
最後に、熱発生と免疫応答の制御に関与する重複するシグナル伝達経路を実証する新しいデータを強調します。
We only briefly discuss the neuronal circuitry that drives fever and the evolutionarily conserved heat shock protein (HSP) response (BOX 1), but refer the reader to recent comprehensive reviews for additional information on these topics as well as on the contributions of hypothermia to limiting inflammation.26–30
発熱を引き起こす神経回路と進化的に保存された熱ショックタンパク質 (HSP) 反応については簡単に説明するだけです (ボックス 1)。
ただし、これらのトピックや炎症の抑制に対する低体温療法の貢献に関する追加情報については、最近の包括的なレビューを参照してください。26–30
以後の解説は、論文をGoogle翻訳でお読みください。
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4786079/