








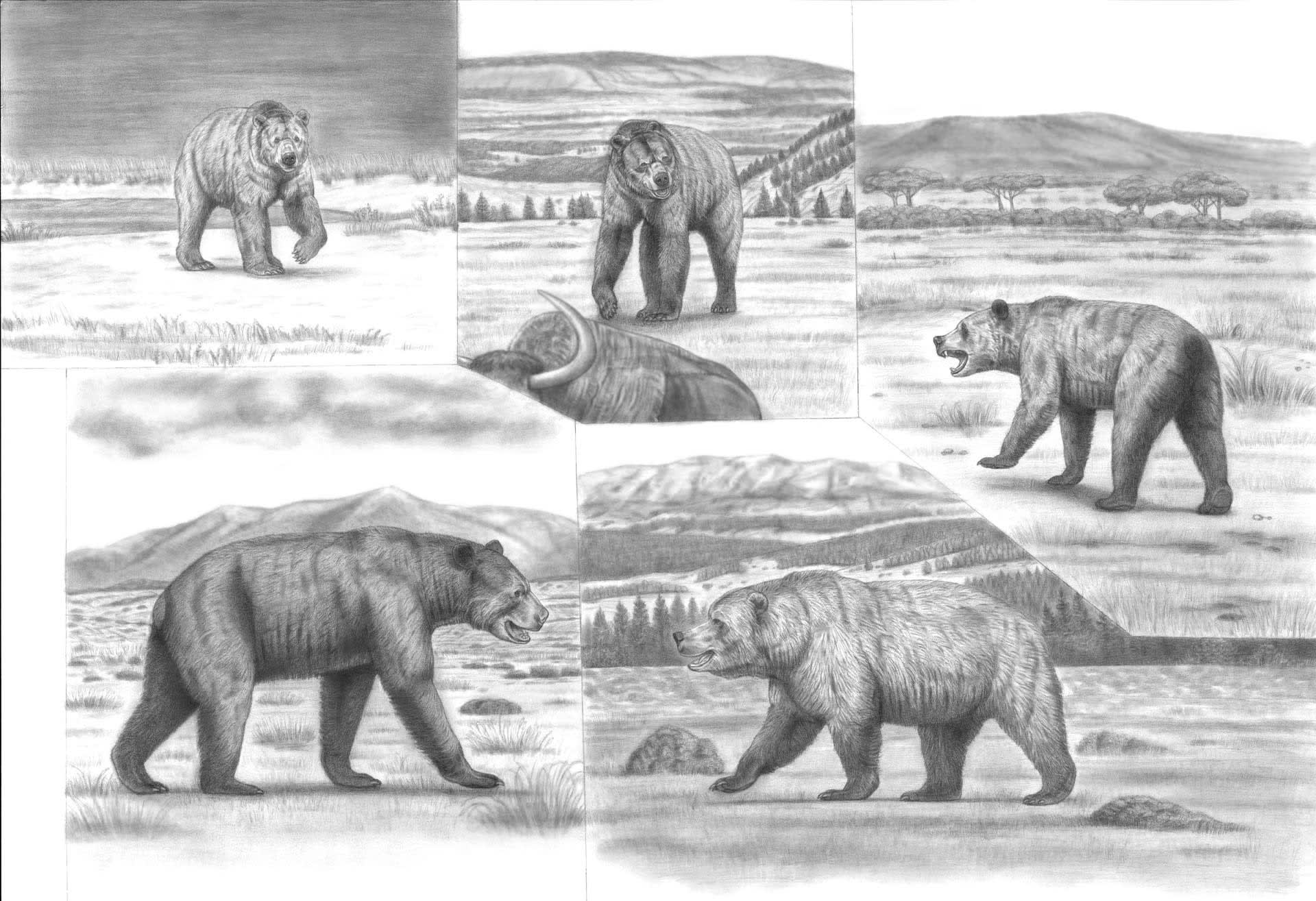
Battle Beyond Epochs 'The Most Colossal Bears of All Time'
クマ科は、食肉目全体の進化史の中でも、最も大きく力強い種類を輩出してきたグループです。彼らの食生の多様さが一因と考えられますが、サイズの大型化の度合いにおいて他の食肉類を圧倒する凄さがあります。
そんなクマ科の中でも指折りの大きさの化石種を選別し、例によって時空を超えた顔合わせを企図した旧作の、当時よりも高画質、大判での再アップ。
無論、生息年代も分布域も異なっていたので、実際にこれらがinteractすることはあり得なかったわけです。想像するのも恐ろしいですが、平均体重が現生のホッキョクグマとコディアックヒグマのそれを凌いだほどの強大な古代熊がもし一堂に会したなら、どんな事態になるのか…

(超大判オリジナルサイズ画像(高画質)) All images by ©the Saber Panther(All rights reserved)
イラスト ©the Saber Panther (All rights reserved)
●Species
上段左から時計回りに:
タイラントグマ(更新世のヒグマないしホッキョククマ、あるいは前者から後者へ移行する過渡期段階のクマ。正式の呼び名ではありません)
Ursus (arctos / maritimus) tyrannus
(更新世後期・北極圏)
キタアメリカジャイアントショートフェイスベア
Arctodus simus yukonensis
(更新世後期・北米)
アフリカショートフェイスベア
Agriotherium africanum
(中新世-鮮新世アフリカ)
ガムスルツェン・ホラアナグマ
Ursus ingressus
(更新世後期・ヨーロッパ)
Arctotheium angustidens
(更新世中期・南米)
[アイスエイジ・ベアーズ]
ヴュルム氷期のブリテンは周氷河地形に分類され、南部地域であっても、現在のアラスカはハドソン湾周辺を彷彿させる、荒涼たる土地が拡がっていたと考えられます。
ホッキョクグマの分布していた形跡すらも-もっとも、尺骨一本ですが-、知られているのです。
ロンドンのキューブリッジ近辺で発掘されたこの尺骨は、遠位部骨端の欠損を補修した上での直径が、48.5cm になります。アルクトテリウム angustidens(クマ科最大種)の最大個体のもの(同57cm)に比べて、長さ、骨幅ともに大きく劣るものですが、Kurten(1964) はホッキョクグマの亜成獣個体の骨だと考えました。
ホッキョクグマは若い種であり、ヒグマと分化した時期はおよそ10万年前と推測されていますが、個人的には、ヴュルム期のホッキョクグマの古亜種にヒグマ的形質が残存していた可能性をこそ、考慮すべきだと思います。
もうひとつ別のクマ科大型種、ホラアナグマ(Ursus spelaeus)も、その分布の北端にあたる地域として、エム間氷期、ヴュルム氷期にはブリテン南部にまで進出していました。
ユーラシアの三大アイスエイジ・ベアーズの分布が重複する、唯一の場所がブリテンであったと言えます。
ウーリーマンモスやケナガサイの一大産地としても知られるブリテンは、荒涼殺伐たる西の最果てというイメージと裏腹に、氷河期を最もよく象徴するメガファウナ(ウーリーマンモス、ケナガサイ、ホラアナグマ、オオツノジカ…)の豊穣さに、特徴づけられた場所でもあったのです。
なお、Kurten(1964) が提起したるこのクマの学名、Ursus maritimus tyrannus の tyrannus の英訳、'tyrant'(暴君)を頭に付けて、かつホッキョクグマかヒグマかの分類が厳密には未定である事情を鑑み、「タイラントグマ」なる俗称を試みに当てましたが、正式な名称ではありません。
[ジャイアントショートフェイスベアの食性 : 最強クラスの大型食肉類]
メガネグマ亜科ショートフェイスベア群に関する最近の複数の形態測定学的、機能形態学的分析(Sorkin, 2006, Figueirido et. al., 2009, Figueirido et. al., 2010) は、いずれもショートフェイスベアの食性は雑食であったこと、肉の消費は主に、kleptoparasitism、すなわちスカヴェンジングに拠っていたという仮説を提出しています。
古くKurten(1967)の主張に起源を持つショートフェイスベア群がハイパーカーニヴォラであったとする説は、現在では支持されなくなっているようです。実際には、既にEmslie & Czaplewski(1985)が比較解剖学的分析から、雑食性であったとする仮説を出していました。
やはり最近の分子系統学の成果(Mitchell et. al., 2016)から、北米のアルクトドゥス属は南米のショートフェイスベア群(アルクトテリウム属とメガネグマ属)から、推測されていたより早期(およそ480万年前)に分岐していたらしいことが判明しているのですが、それでも大型種について言えば、南北アメリカのショートフェイスベア群の形態は非常に類似しています。
食肉目史上最大級のサイズ、忍び寄り型、追跡走行型のいずれの狩猟にも比較的適応性が乏しい(Sorkin, 2006)とされるポストクラニアル形態、比較的吻部が短く※強大な顎力を有する頭骨など、ジャイアントショートフェイスベアの特異な形態要素についても、最新の食性の仮説に照らしてみれば、より良く解明されると言われています。もちろん、雑食性の仮説自体は、主に頭蓋‐歯形(cranio-dental)の形質分析(この点では特に、Sorkin, 2006 の研究が包括的で、徹底している)に基づいていることは、言うまでもありません。
(※比較的短吻 'relatively short snouted' というのは曖昧な表現ですが、現生メガネグマを含むショートフェイスベア群が、実はそれほど短吻ではなかったということを、他のクマ科種との比較解剖学的分析を通じて例示した "Demythologizing Arctodus simus, the 'short-faced' long-legged and predaceous bear that never was" Figueirido et. al.,2010 といった研究もあります。参考まで)
恐らく、ジャイアントショートフェイスベアは時に大物猟も行っていたことでしょうが、Figueirido et. al., 2010に拠ると、その生態は現生の北米内陸部のヒグマの一部にみられるものと、似通っていただろうということです。しかし現生ヒグマに比べて、クレプトパラサイティズムにより重きが置かれていたということなのでしょう。
ジャイアントショートフェイスベア群のように巨大な肉食獣が、摂食の少なからぬ割合をスカヴェンジングに拠っていたという説は信じ難い印象も受けますが、更新世のアメリカ大陸において、消費できる大型動物の死骸数の規模は現代より遥かに大きかったとされています。剣歯猫が共存していた地域にてその傾向は特に顕著であったはずで、極度に特化したハイパーカーニヴォラであった剣歯猫は、その歯の構造上、主に獲物の柔らかい部位の肉しか消費できず、それ以外は消費されずに残されていたと考えられるのです(ショートフェイスベア群は、骨を砕くことのできる歯形をしていた)。
巨体と、プロポーション的に伸長した前肢(Sorkin, 2006)で広大な領域をカヴァーし、時に狩りもし、時に他の大型肉食獣を追い払って獲物を奪取する(つまり、クレプトパラサイティズムにも適応している)、非常にヴァーサタイルな雑食獣の姿が、浮かび上がってきます。ある意味「最強」と言えそうな、これほど適応力にすぐれた強大な肉食獣が、なぜ絶滅に追いやられてしまったのか、逆に不思議に思えてくるほどです。
ついでながら付け加えると、中新世から鮮新世にかけてのアグリオテリウム属(ウルサヴス亜科)も比較的に短吻とされ、メガネグマ亜科と系統的に縁遠いにもかかわらず(もっとも、アグリオテリウム属がメガネグマ亜科の祖先筋であった可能性も否定はできないと思います)ショートフェイスベアの通称で呼ばれていますが、アグリオテウムの食性については何とも断定し難い状態です。Sorkin(2006)はアグリオテリウム属種の頭骨もメガネグマ亜科の種類と共に「ショートフェイスベアのモーフォタイプ」として分析サンプルに含めて、どちらの食性も雑食であったとする仮説を出しているのですが、アグリオテリウム africanum(アフリカ産の最大種)の頭蓋-歯形の機能形態学的分析に取り組んだWroe et. al.(2013)の研究は、同種の草食の可能性を除外しないまでも、それが摂食に占める割合は小さかったことを示唆しているからです。

[アフリカショートフェイスベア(アグリオテリウム属最大種)について]
とまれ、最後に付け加えると、アフリカショートフェイスベアは類稀な顎の力の持ち主でした。肉食獣の咬筋力の一連の研究で著名なWroe(2013)によると、肉歯類やエンテロドン科種を含まずに食肉類のみに限定した場合、彼が分析に携わった中で咬筋力最強はアフリカショートフェイスベアだったということです。
更新世後期の動物相を代表する一種、「ホラアナグマ」の分類について、古遺伝学の分野の最新研究が一致して提示する仮説は、Ursus spelaeus として一括りにされてきた古代熊の中に、実は複数(最新仮説では4種)の異なるハプロタイプが認められるというものです(Orland et al., 2002, Hofreiter et al., 2004, Stiller et al., 2009)。形態測定学、放射線年代学各分野の近年の研究成果も、複数種説を補強するでしょう。
オーストリアのガムスルツェン洞窟で見つかるクマは、特に他との遺伝的、さらには形質的差異も顕著であり、現在では学名 Ursus ingressus 、通称「ガムスルツェン・ホラアナグマ Gamssulzen Cave bear」という固有種としての分類が定着しているのです(Hofreiter et al., 2004, Stiller et al., 2009)。他の三つのハプロタイプ間にも形態やサイズの違いは存在するとされますが、いずれもUrsus spelaeusの亜種として分類する説が主流の模様(Rabeder et al., 2004)。
このうち、ホラアナグマ系統の最大種であるガムスルツェン・ホラアナグマ(Ursus ingressus)と、最小亜種とされラメッシュ洞窟で主に見つかるラメッシュ・ホラアナグマ(Ursus spelaeus eremus)は、更新世後期の一時、具体的には1万5千年ほどの間、オーストリアの一部地域で共存していたらしいことが判っています(Hofreiter et al.,2004)。比較的長期間、分布域がほぼ重複していたにもかかわらず、2種間の遺伝的差異は保たれていたということです。
現在でもユーラシアや北米大陸にて、ヒグマとクロクマなど、複数の異なるクマ科種の共存の例が知られますが、いずれも雑食性の種類で食性の比重を柔軟に変化させられることが、スムーズな共生を可能にする要因であると考えられます。他方、ホラアナグマ系統はいずれも、歯形や骨の同位体分析からほぼ完全な草食性であったことが判っており、異なる種類の「草食性のクマ」の共存例は、現代には存在しません。
この点で、オーストリア産ホラアナグマ系統2種間の棲み分けの実態について、主に骨の同位体分析を通して解明に当たった研究、'Niche partitioning between two sympatric genetically distinct cave bears(Ursus spelaeus and Ursus ingressus and brown bear(Ursus arctos) from Austria : Isotopic evidence from fossil bones', Bocherens et al., 2011
は、興味深いと思います。
著者によると、ガムスルツェン・ホラアナグマとラメッシュ・ホラアナグマはそれぞれ異なる種類の植物を主に摂食していたらしいことと、酸素同位体分析から異なる水源の水の利用が示唆されることから、厳密には棲み処が分かたれていたことが推測されますが、2種の分布は極めて狭い(人間の徒歩で数時間で移動が可能な)範囲に集中していました。
この場所的棲み分けが、短期的な寒冷、温暖といった気候変動に合わせて生じた時間的棲み分けでもあったのかどうか、判然としないようですが、そうではなかったとすると、ホラアナグマの系統が実に極めて強固な縄張り性(territoriality)を有し、種間の遺伝子流動が防がれていたことが窺い知れます。
なお、ガムスルツェン・ホラアナグマは寒冷期のクマで、寒地適応の結果、比較的温暖期に繫栄したであろうラメッシュ・ホラアナグマよりも大型化したという仮説も見受けられます。実際、ホラアナグマが一種のみと認識されていた時代のクルテンなど、間氷期、氷期の違いでサイズに著しい多様性が生じていたことを指摘していました。個人的に、ホラアナグマ系統各種間のサイズ差の原因についても、より詳しい説明を望むところです。









