最近のDNA検討から、イヌは日本列島から生まれたとする説が出ているようだ
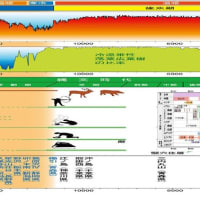
旧石器時代後期から土器発生までの時期
尖頭器から石鏃開発以前、それは有舌尖頭器の時期に当たる
有舌尖頭器 投げ槍

有茎石鏃
大型動物が絶えて、狩りの形が変わり、小動物の狩りに変わる前
この時期にイヌが家畜として飼われることになり、イヌとの共同の狩りで
槍とされる大型尖頭器よりも投げ槍とされる有舌尖頭器の全盛期となるのでは無いか

旧石器後期から縄文時代に入る前、イヌがすでに家畜として飼われるようになり、槍での狩りではイヌの利用は効果的でなかったが、投げ槍を使うようになるとイヌを利用した狩りは、好都合だったのではないか、投げ槍では危険が生じるので、イヌであればそれは避けられることになる。投げ槍を使うようになり、イヌを使い、狩りが効果的に行えるようになっていたのではないか

有舌尖頭器、有茎尖頭器と呼ばれている 7センチほどの長さ
有舌尖頭器は次第に小さくなり、石鏃との境界は分らないようだ。茎、舌が消え更に軽くなったものは石鏃として良いのでは、茎が有るものは未だ投げ槍の範疇に入るのだろうか。この時期に土器で隆起線文、爪形文の土器が造られこれが列島全体に行き渡り、土器の造られる量も格段に増加しているようだ。
これが早くから行えたのは日本列島の植生など特性であるのでは
定住に近い生業も可能となって来ていたのでは無いか、そして有茎尖頭器の発展する時期に、土器の発生となるのでは。

図はお借りしました
引用します----ーーーーーーーーーーーー
動物相の変化と尖頭器の消長
更新世末から完新世初頭にかけては、日本のみならず北アメリカ大陸・アジア大陸においても尖頭器の発達が著しい。これらの地域ではマンモスやバイソンなど洪積世の寒冷気候を好んだ大形獣を対象とする狩猟具が求められ、投げ槍の槍先として用いられた各種尖頭器の出現をみた。
日本においても、当初は大形獣を対象とした狩猟具として生まれたが、洪積世末期に海進によって大陸から切り離され、大形獣の絶滅が早かったという特殊な条件が加わって、イノシシ・ニホンジカなどが主な狩猟対象となった。これらの、嗅覚が鋭く行動の機敏な動物の捕獲には、手持ちの槍よりも投げ槍が狩猟具として適していたものと考えられ、特に有舌尖頭器の急増は、こうした事情を物語っていると推定される。
やがて弓矢の発明とともに、タヌキやウサギなどの小動物も狩猟対象となっていった。そして、弓矢と槍の中間的な機能を果たした投げ槍(槍先形尖頭器)は弓矢の普及によって消滅していく。
一方で、縄文時代前期以降は採集・漁撈の充実および定住生活のいっそうの進展とともに落とし穴を利用する待ち伏せ狩猟も増加していくのである。
++++そうなのか イヌとの関係は
----ーーーーーーーーーーーー
旧石器時代から縄文時代へ きゅうせっきじだいからじょうもんじだいへ
約1万5千年前から1万年前の日本列島には、細石刃、有舌尖頭器、大型の石斧(神子柴型石斧)を主体とする三つの石器群がめまぐるしく登場します。細石刃とは長さが2~3cmのたいへん小さな石刃で、細長い溝のきられた骨製の柄に何個もはめ込まれて、投げ槍のような便われ方をしたと考えらています。富山県では小矢部市の日の宮遺跡で細石刃を剥がすための石核が出土しています。
有舌尖頭器とは柄を装着するために舌状に作り出された5~l0cmの長さの槍先です。
日本で最も古い土器は、この石器が盛んに使用されていた頃に出現しています。
神子柴型石斧とは短冊形で長さが約20cm程度と大きく、刃部の磨き上げられた重量感のある石斧で、木材の伐採に用いられたと考えられています。この石器には神子柴型尖頭器とよばれるスリムで長大な槍先や特殊な砥石などが伴っており、丸底で線状の模様がつけられた隆線文土器もみられます。富山県では、大沢野町の野沢遺跡や八木山大野遺跡で神子柴型石斧が出土しています。
気候の温暖化や狩猟対象の大型から小型の哺乳動物への変化といった条件が、道具の変化をもたらし、縄文文化への橋渡しを行ったと考えられます。
白岩尾掛遺跡から採集された有舌尖頭器(上段左側)
富山県の最古の土器(下段3点) 野沢遺跡から出土した神子柴型石斧(長さ23.2cm、重さ485g)
お問い合わせ
所属課室:教育委員会生涯学習・文化財室
〒930-8501 富山市新総曲輪1-7 県庁南別館4階
----ーーーーーーーーーーーー
学位論文要旨 No 123911
著者(漢字) 長井,謙治
著者(英字) 著者(カナ) ナガイ,ケンジ
標題(和) 日本列島における有舌尖頭器の研究 : 実験考古学の実践的研究
標題(洋)
報告番号 123911
報告番号 甲23911
学位授与日 2008.03.24
学位種別 課程博士
学位種類 博士(環境学)
学位記番号 博創域第377号
研究科 新領域創成科学研究科
専攻 社会文化環境学専攻
論文審査委員 主査: 東京大学 教授 佐藤,宏之
東京大学 教授 辻,誠一郎
東京大学 教授 鬼頭,秀一
東京大学 准教授 清水,亮
東京大学 准教授 清家,剛
内容要旨 要旨を表示する
有舌尖頭器とは更新世/完新世移行期に作られた狩猟用石器である。
近年、日本列島の更新世末期における陸上生態系/植生史が明らかとなることで、針葉樹林から温帯の落葉広葉樹林へという陸上生態系の急変が明らかとなっている。この急変とほとんど軸を一にして、日本列島に出現/消滅した狩猟具のひとつが有舌尖頭器である。旧石器文化から縄文文化へという文化の変化と画期に消長関係が合致する有舌尖頭器は、縄文文化形成の動態を解くきわめて重要な考古文物としてこれまでに学界で注目されてきた。
同石器は、晩氷期中盤から末期にかけて地域差をもちつつ、日本列島(九州―四国―本州をひとつの陸塊島とし、北海道を大陸から突き出した半島とする古地理)のほぼ全域で盛行する。晩氷期後半に凡そ対応する約13,600 y.B.P.から9,800 y.B.P頃における日本列島の有舌尖頭器群が本論文のおもな検討対象である。
これまでの研究は「かたち」の文化規範を探ることに端を発し、その延長上にある諸研究は現在の資料から矛盾をきたしている。第1章では一系統的起源論を前提とした「形」・「型」の研究史を回顧して、有舌尖頭器の学史にある上記の問題を詳しく描出する。
第2章では石器づくりの実験考古学により、独自の方法論を生みだす。まずは有舌尖頭器の復元製作を行い、その複製資料を使って剥離面の解析を行い、石器作り動作の型を指摘する。使用するデータは申請者・T.Dillardの複製品、民族資料等である。
結論として、石器作りには「正位」・「逆位」の加工法があり、この2つの加工法は共に辺に対して水平の剥離を生み出すものの、辺に対して上下に傾く剥離をより安定して生み出す傾向にあることを明らかとする。この結論は複製資料の剥離軸を定量的に調べることで導出された。すなわち、右辺で左下方へ、左辺で右上方へと傾く右肩上がりの剥離面は正位の剥離により効率的につくられ、右辺で左上方へ、左辺で右下方へと傾く左肩上がりの剥離面は逆位の剥離により効率的につくられる傾向が明らかとなった。これは石器作りの動作とそれが生みだす痕跡の間にある強い相関関係として指摘された。第3章以降は、このような傾く剥離をもつ有舌尖頭器を対象とした考古学的分析をおこなった。
第3章では、斜行有舌尖頭器を提唱して、その日本列島における分布と展開を検討した。筆者および先学が集成・紹介した本州・四国・九州・北海道に於ける305遺跡以上の有舌尖頭器が主たる検討の対象となった。
第3章の結論としては、日本列島における斜行有舌尖頭器は本州西半部と北海道に安定して認められ、北海道で左肩上がりの有舌尖頭器を主体とし、本州以西で右肩上がりの有舌尖頭器を主体とすることが指摘された。
日本列島における斜行有舌尖頭器は、北東北地方付近を境とする西と東で斜方向を真逆としたことが明らかとなった。
では、この斜方向の違いは何を物語っているのか。そして、その歴史的意義はどのように解かれるのか。斜方向/剥離順序から石器扱いを読み解いたのが第4章である。
第4章では、二つの石器扱いを導出する。この作業により、日本列島の有舌尖頭器が一系統的存在であるということに対する最初の否定的根拠が提出されることとなった。
二つの石器扱いは石器作りの経験から以下のように導き出された。正位の剥離により連続的な剥離をうみだすとき、左辺で向こうから手前に剥離が進行し(↓)、右辺で手前から向こうに剥離が進行する(↑)。一方、逆位の剥離はすべてこの反対の仕組みとなる。逆位の剥離が手前への剥離である以上、向こうに高所があり、手前に低所があることが打面の強度と剥離誘導の安定性を保障するうえで必要となる。したがって、逆位では右辺で向こうから手前に剥離が進行し(↓)、左辺で手前から向こうへと剥離が進行する(↑)。すなわち、正位の剥離のとき、左辺で向こうから手前(↓)/右辺で手前から向こう(↑)の剥離順序が導出され、逆位の剥離のとき、右辺で向こうから手前(↓)/左辺で手前から向こう(↑)の剥離順序が導出されることを指摘した。(註)向こう、手前などの表現は割り手から見た空間を指す。
以上を踏まえたうえで、考古資料の石器扱いが遺物の詳細な観察により検討された。その結果、正位の石器扱いの証拠を突きつける剥離の順序/向きは、本州・四国の右肩上がりの有舌尖頭器・尖頭器群から導出された。
一方、逆位の石器扱いの証拠を突きつける剥離の順序/向きは、北海道の左肩上がりの有舌尖頭器・尖頭器群から導出された。このように斜方向と加工手順という実証的な手がかりから日本列島における二つの石器扱いが導き出された。
以上の分析を介して重要となったのは、有舌尖頭器の石器扱いが本州・四国と北海道では全く異なるという推論であった。
石器扱いは石器作りの動作により生まれるが、これは学習、伝承されるものであるから、石器扱いの違いとは、各地方ですでに存在する石器作りシステムの違いを反映している可能性が高いと予想された。そこで次に、有舌尖頭器のみならず、斜めの剥離をもつ石器群(斜行石器群)の検討をしたのが第5章以降である。
日本列島の東と西で認められる石器扱いの東西差は、いつの時期、どの程度の期間に、どのような背景から形成されたのか。第5・6章では、日本列島各地における有舌尖頭器の消長関係を明らかにすると共に、東西・2つの有舌尖頭器群を内包する斜行石器群の様相を検討した。東西・斜行石器群の展開:その時間と構造/有舌尖頭器の由来までを検討した。
第3章から第6章までの検討を通して、東西・斜行有舌尖頭器の展開・構造・時間・由来に関する推論は次のように果たされた。
1)日本列島における一方向型の斜状平行剥離を施す有舌尖頭器は、斜方向の差で異なる分布を示す。本州西半部に分布する斜行有舌尖頭器は、その斜方向が右肩上がりとなるが、北海道に分布する斜行有舌尖頭器は左肩上がりとなる。日本列島における斜行有舌尖頭器は、北東北地方付近を境とする西方と東方で斜方向を真逆とする。そしてこの東西差は晩氷期の約13,000 y.B.P.〈約15,500 cal BP〉から約11,000 y.B.P.〈約13,000 cal BP〉頃の日本列島で一時的に併存した。
2)晩氷期・石器群からは石器扱いの差が導出される。そしてこの石器扱いの違いは、石器作りシステムの違いにより顕現化している。
RL-斜行石器群が九州-四国-本州的規模で地域色を有しつつ連なるホライズンを形成しており、
LR-斜行石器群が北海道で固有のホライズンを形成する。
右肩上がりの有舌尖頭器は晩氷期後半のRL-斜行圏(九州-四国-本州)に消長し、左肩上がりの有舌尖頭器は晩氷期のLR-斜行圏(北海道)に消長する。
右肩上がりの有舌尖頭器と左肩上がりの有舌尖頭器は異なる石器作りシステムの中に生まれて消えた。
3)剥離の向きと型式から有舌尖頭器の一系統的起源論を再確認すると、本州以西の小瀬が沢系有舌尖頭器と北海道の立川系有舌尖頭器に右肩上がりないし左肩上がりの有舌尖頭器が認められる。
この場合、立川系から小瀬が沢系の型式変化の間に石器扱いの変化を認めざるを得ず、よって立川系と小瀬が沢系は別系統とみるべき結論に至る。この推論により由来を同じくしない有舌尖頭器の存在が明示的となった。
最後に「先史考古学における動作連鎖論」から東西差の意義を検討した。
北海道・斜行石器群を構成する後半期細石刃石器群からは、強固なシェーンに逆位の石器扱いが組み込まれた内鎖(インサイド・オブ・シェーン)および外鎖(アウト・オブ・シェーン)を形成する石器作り(身体技法)が導き出された。北海道・左肩上がりの有舌尖頭器は北海道・固有の動作連鎖により作られたと考えられる。
他方、本州以西の西の世界には北海道と同型の動作連鎖は認められないと考えられる。このように、本州以西と北海道が互いに異なる石器作りの文化的伝統を有しており、それぞれ異なる石器作りの文化的伝統から二つの有舌尖頭器が作られていたことを明らかとした。
これまでに、我が国の有舌尖頭器はごく一般的に大陸から渡来したと見なされてきた。
すなわち、旧石器時代から縄文時代へという文化の変化は、大陸起源という新たな文化的接触にその形成の契機を求めてきた。北海道を伝播経路とし、そして、本土へと渡来した大陸起源の文物は、我が国・当地の文化と融合・融和し、所与の縄文文化を形成した。こうした理解を支えたのがまさに日本(北海道・本州・四国・九州)におけるひとつの有舌尖頭器文化であった。
ところが、日本列島の有舌尖頭器は、東西の異なる石器作り文化に生まれて消えた結論に達する。
この結論は同時に、日本の一系統的有舌尖頭器が成立する余地が無いことを実証的に説明できたことを意味するのであり、所与の縄文文化を否定するものであった。
このように、本研究は石器づくりの実験考古学による独自の分析手法を介して、晩氷期の日本列島における2つの石器文化を描き出すことで、日本の一系統的有舌尖頭器が成立する余地が無いことを実証的に説明している。
----ーーーーーーーーーーーー
●ユーラシア東部のイヌへのニホンオオカミからの遺伝的寄与
上述のニホンオオカミとユーラシア東部系統のイヌとの間の遺伝子流動の問題は、以下の4点にわたります。(1)ニホンオオカミとイヌとの間の交雑の年代と場所です。(2)ニホンオオカミとイヌとの間の遺伝子流動(交配と戻し交雑)が、単方向だったのか、そうだとしてどちら側からなのか、また、あるいは双方向の遺伝子流動があったのか、ということです。(3)遺伝子流動があったとしたら、どの程度だったのか、ということです。(4)遺伝子流動の回数です。
まず(1)については、古代ゲノムデータの解析にわり、ニホンオオカミのゲノムの一部領域は9500年前頃にシベリアにいたイヌのゲノムにすでに含まれていた、と明らかになっています(Gojobori et al., 2021)。つまり、ニホンオオカミとイヌとの間の交雑は少なくとも9500年以上前に起きていた、と推定できます。日本列島で最古のイヌの骨は9600年前頃と報告されており、日本列島にイヌがヒトとともに到来したのはその少し前と推測できます。こうした知見から、ニホンオオカミの祖先とイヌとの間の交雑はアジア東部において1万年以上前に起き、すでにニホンオオカミに由来するゲノムの一部領域を有していたイヌが縄文時代に日本列島に到来した、と考えられます。
(2)査読前論文(Gojobori et al., 2021)では、ニホンオオカミからユーラシア東部のイヌへの遺伝子流動が示されています。他のハイイロオオカミ集団とイヌとの間の遺伝子流動の方向はイヌからハイイロオオカミなので(関連記事)、その逆というわけです。
(3)ニホンオオカミのゲノムを最も多く有しているのはディンゴとNGSD で、最大で5.5%程度です(図3)。次いで多いのは秋田犬や紀州犬や柴犬などの日本犬とアジア東部および東部の在来イヌです(図3)。一方、ヨーロッパのイヌやアフリカのイヌからは有意なニホンオオカミからの遺伝子流動は検出されていません。
(4)世界の犬種の遺伝的類似性をユーラシア東部の古い系統であるディンゴ/ NGSDとニホンオオカミについてそれぞれ見ると、ディンゴ/ NGSDに遺伝的に近いイヌはニホンオオカミにも近く、ディンゴ/ NGSDから遠いイヌはニホンオオカミからも遠い傾向があります(Gojobori et al., 2021)。こうした傾向は、ユーラシア東部のイヌの祖先でニホンオオカミからの遺伝子流動により生じた、と推定されています。そのため、ニホンオオカミからイヌへの遺伝子流動は1 回だけ起きた、と推測されます。これらの関係は図4で示されています。以下は本論文の図4です。
画像
ニホンオオカミを除くユーラシア大陸のハイイロオオカミ集団とイヌの系統が分岐した年代は4万~2万年前頃で、次にイヌの東西ユーラシア系統の分岐は24000~17000年前頃と推定されています(関連記事)。ニホンオオカミはユーラシア大陸のハイイロオオカミ集団から分岐しており、イヌの東西ユーラシア系統の分岐前にイヌ系統から分岐しているので、ニホンオオカミの系統の分岐は40000~17000年前頃と推定できます。ニホンオオカミ系統は、イヌの東西ユーラシア系統が分岐した後に、ユーラシア東部系統の祖先と交雑した、と考えられます。
ニホンオオカミとイヌとの間の交雑については、実際に起きていた証拠が報告されています。上述のように、シーボルトが江戸時代にオランダに送った標本に、ヤマイヌと呼ばれている個体がありました。この個体由来のDNA を詳しく調べると、ニホンオオカミのゲノムにイヌのゲノムが入っている個体と分かりました。このヤマイヌ個体にゲノムを提供した個体に最も近いと推定されるのは現在の柴犬で、39%ものゲノム領域がこのヤマイヌ個体に含まれていました(Gojobori et al., 2021)。
本論文は憶測と断りつつ、もし江戸時代に生きていた日本犬のゲノムと比較できるならば、このヤマイヌ個体に含まれるイヌゲノムの割合はもっと高い値、おそらくは50%程度になるのではないか、と推測しています。仮に50%程度ならば、このヤマイヌ個体はイヌとニホンオオカミとの交雑第一世代の個体でと考えられます。また、このヤマイヌ個体からmtDNAが調べられており、この個体のmtDNAはニホンオオカミ型と報告されています。mtDNAは母系遺伝なので、このヤマイヌ個体は、母親がニホンオオカミで、父親がイヌと推定できます。
●ニホンオオカミの歴史
以上の知見を総合すると、ニホンオオカミは40000~17000年前頃にユーラシアのハイイロオオカミ集団から分岐した、ユーラシア大陸東部に分布していたハイイロオオカミの1系統で、最後まで生き残った場所が日本列島南部(本州と四国と九州)でした。ニホンオオカミ系統はアジア東部大陸部に分布していた頃にはユーラシア東部系統のイヌの祖先と交雑し、現在もユーラシア東部やオセアニアのイヌのゲノムには、広くニホンオオカミ由来のゲノム領域が存在します。他のハイイロオオカミ集団とは異なるニホンオオカミの特徴は、イヌとの交雑後に交雑個体がイヌに戻し交配していることです。
ニホンオオカミがイヌと最も近縁なハイイロオオカミの系統で、さらにユーラシア東部のイヌのゲノムにニホンオオカミに由来するゲノム領域が含まれていることから、ニホンオオカミの系統はイヌの家畜化に大きく関わってきた可能性が高そうです。こうした独自のハイイロオオカミの1系統が100年前頃まで生き残っていたことは驚くべきである、と本論文は評価します。ただ、ユーラシア大陸から隔離された日本列島であったからこそニホンオオカミが生き残れた、とも言えそうです。本論文は、このようなニホンオオカミの重要性から、すでに絶滅してしまったことを残念に思う、と述べています。
図はお借りしました